-
- Figure 1
-
- Figure 2
-
- Figure 3
-
- Figure 4
-
- Figure 5
-
- Figure 6
-
- Figure 7
-
- Figure 8
-
- Figure 9
-
- Figure 10
-
- Figure 11
-
- Figure 12
-
- Figure 13
-
- Figure 14
-
- Figure 15
-
- Figure 16
-
- Figure 17
-
- Figure 18
-
- Figure 19
The Kidney
Microscopic Structure ![]()
The kidney is composed of stroma and parenchyma.
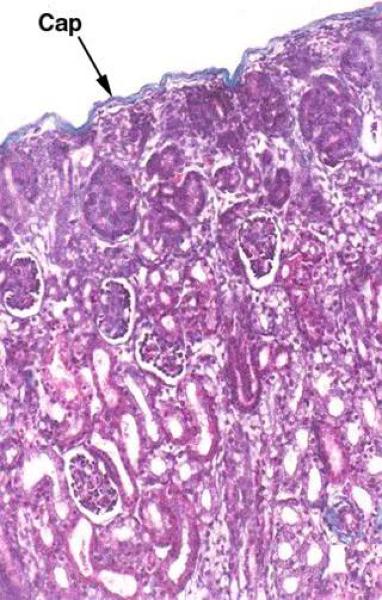
The stroma is represented by outer capsule and reduced reticular scaffolding.
Unlike the capsule of many other organs, this one doesn’t send septa down into the parenchyma.
There is almost no CT scaffolding in the kidney’s internal volume.
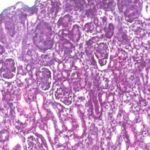
At low magnification, The parenchyma is differentiated into an outer cortex and an inner medulla.
The renal cortex comprises two distinct regions: the cortex proper or cortical labyrinth and the medullary rays.
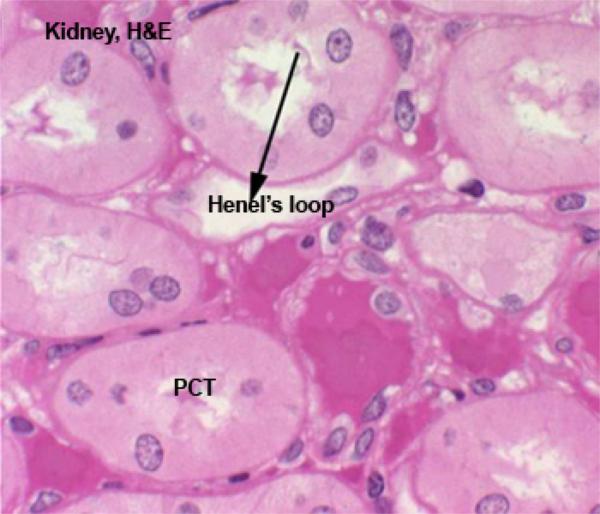
The cortical labyrinth contains renal corpuscles, proximal and distal convoluted tubules and arched collecting ducts.
The medullary rays are formed of descending and ascending limbs of nephron loop and straight collecting tubules.
The medulla is deep to the cortex in it there are no renal corpuscles, only tubules.
Nephron
The nephron is the structural and functional unit of the kidney of which approximately 1.3 millions are present in each kidney.
There are several distinct portions to a nephron.
These are the renal corpuscle (which encloses the glomerular tuft), the proximal convoluted tubule, the loop of Henle, a distal convoluted tubule, and a short collecting tubule.
Each nephron is a continuous tubular unit, emptying via its collecting tubule into a collecting duct of larger size, which receives input from many nephrons.
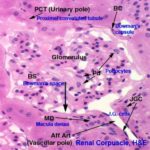
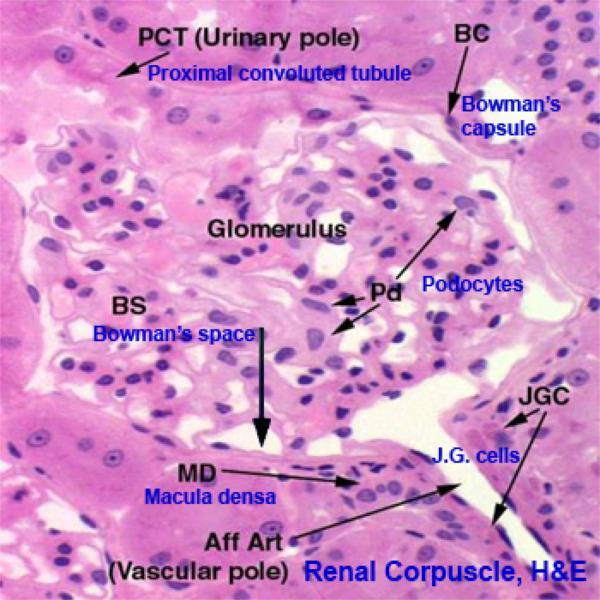
The renal corpuscle is the first part of the nephron.
The corpuscle has two components: 1) the tuft of capillaries constituting the glomerulus; and 2) the outside wall, which creates a well-defined space around the tuft, the Bowman’s space.
The epithelium of the corpuscle has two portions.
The parietal layer, forming the outside wall, is a thin simple squamous type, resting on a basal lamina. This squamous epithelium is continuous with the cuboidal epithelium of the proximal convoluted tubule.
At the point of entry of the afferent arteriole, the visceral layer is continuous with the parietal layer of the capsule, sealing off Bowman’s space.
The visceral layer of the renal corpuscle is a very specialized epithelium: it takes the form of podocytes ![]()
![]()
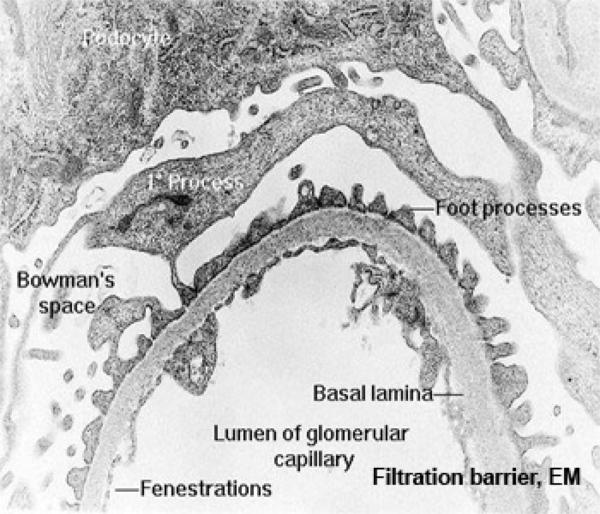
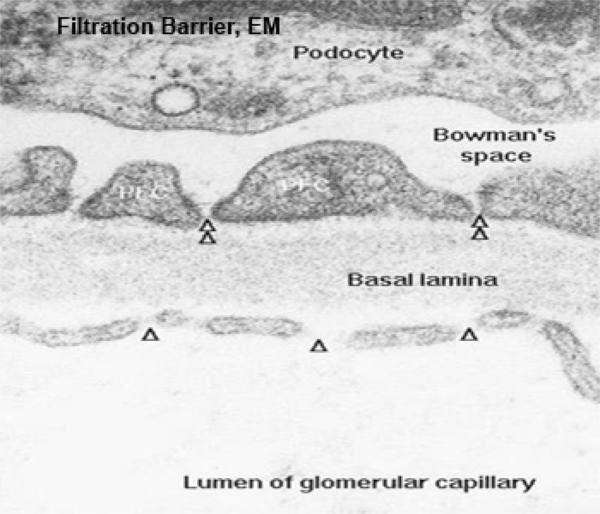
Podocytes have long, extensively arborized foot processes which wrap around the capillaries of the glomerulus.
Podocytes can be seen with the light microscope, but their processes can’t.
The processes cover the entire surface of the capillaries in the glomerulus, except for slits between adjacent processes; these slits are the actual site of filtration.
Glomerulus ![]()
![]()
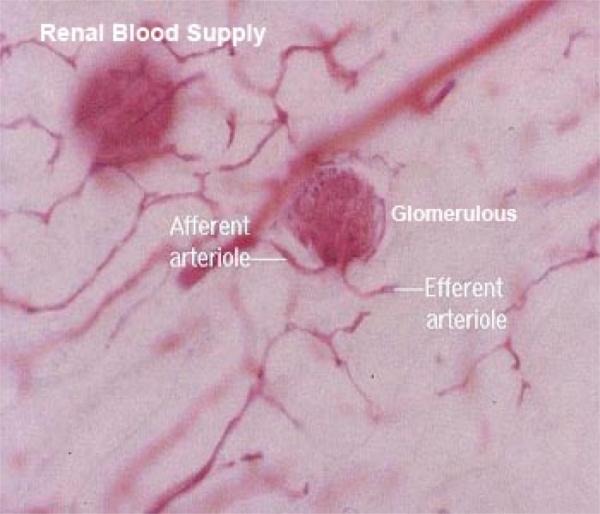
The glomerulus is a tuft of blood capillaries supplied by an afferent arteriole and drained by a smaller efferent arteriole.
The capillary endothelium is fenestrated and rests on a thick basal lamina.
The capillaries are surrounded by mesangial cells.
Blood passing through the capillaries is under some considerable degree of arterial pressure; this causes plasma to be forced out of the capillaries and through the slits between the podocyte foot processes.
It collects in the sealed space of the renal capsule, and flows thence into the proximal convoluted tubule.
Since the slits are far too small to pass formed elements of the blood, these normally should not be present in urine. Furthermore, the thickened basement membrane between the podocyte processes and the capillary walls acts as a “super filter” and removes from the filtrate most molecules over a certain molecular weight.
Thus large proteins shouldn’t be present in normal urine, either.
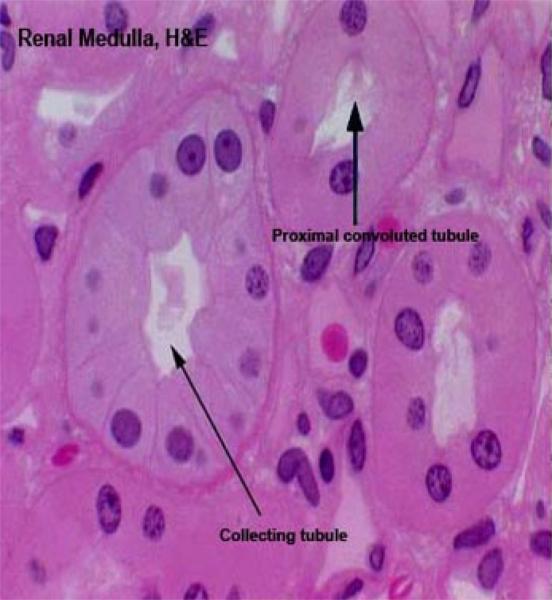
From Bowman’s space, the filtrate (driven by the hydraulic pressure of the glomerular blood flow) passes to the proximal convoluted tubule (PCT).
Here much of it is reabsorbed. Approximately seventy-five percent of glomerular filtrate is reabsorbed from the PCT. If it weren’t, an animal would die quickly of dehydration: the average human filters 1200 liters of blood per day, but passes only about a liter of urine. Obviously most of it has to be reclaimed.
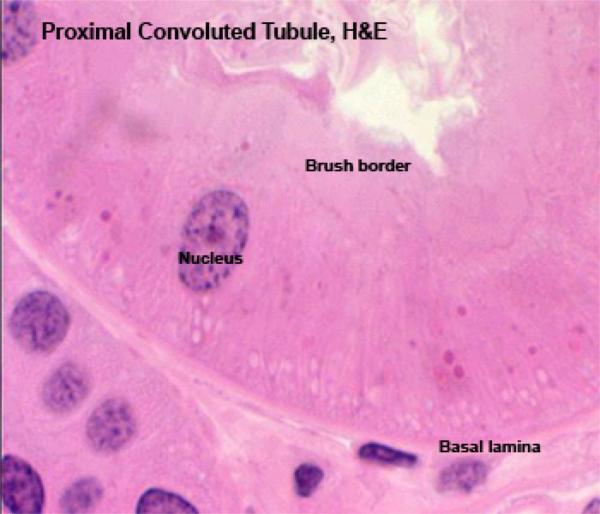
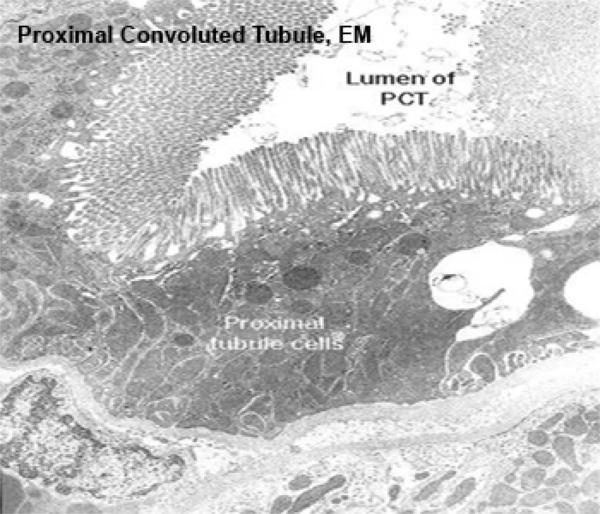
Most of the cortical tissue around the renal corpuscles is proximal tubules.
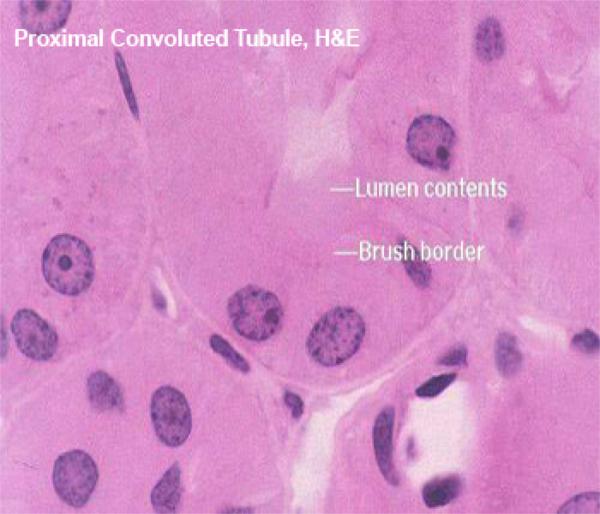
These are made of tall cuboidal epithelium with ill-defined cell boundaries and spherical nuclei located near the base.
The luminal surface of the cells has a distinct and prominent microvillous brush border which almost completely fills the lumen.
The lumina of PCT’s are often collapsed (due to cessation of glomerular blood flow upon death), Only the PCT has such a brush border. The other kidney tubules don’t.
The PCT’s brush border is of course a specialization for increasing surface area for absorptive activity.
The cytoplasm of the PCT epithelial cells stains intensely due to high a high content of organelles, principally mitochondria.
The basal plasma membrane exhibt deep basal infoldings into the cells. These infoldings are closely associated to columns of mitochondria. This mitochondrial arrangement gives rise to the appearance of basal striations in light microscopy.
The bulk of the renal cortex is composed of PCTs since the PCT is the longest and most convoluted part of the nephron. The PCT extensively modifies the original filtrates with respect to concentration and composition: in particular it’s very active in the resorption of salts and water.
About half a pound of glucose and three pounds of sodium chloride per day are recovered in the PCT.
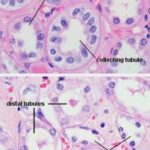
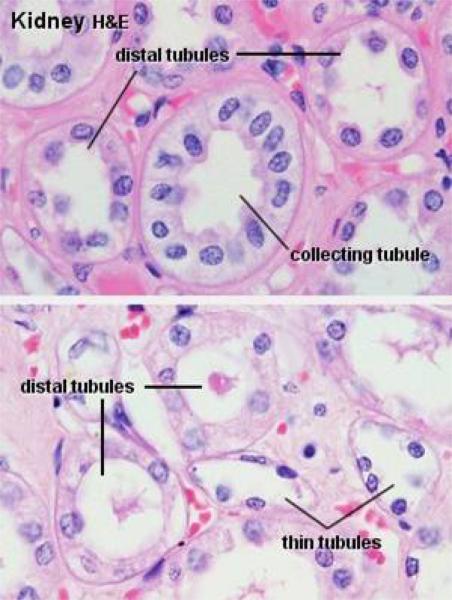
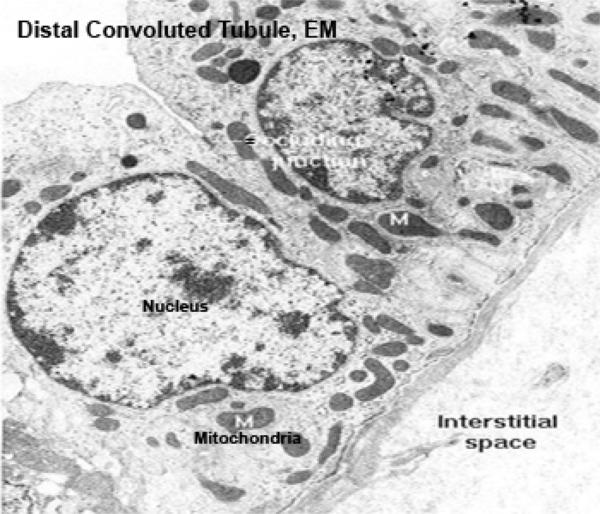
As it comes back up into the cortex region the filtrate passes from the loop of Henle into the distal tubule (DT). The walls of the DT, like those of the PCT, are of simple cuboidal epithelium, but they lack a brush border. They also tend to be larger in cross section than the PCT.
The DT isn’t specialized for absorption. Its role is to pump sodium ions out of its lumen into the surrounding intracellular space. It does this by active transport, using membrane bound pumps.
The DCT may be differentiated from the PCT on the basis of the following characteristic features:
Absence of brush border,
In cross section the DCT has a large more clearly defined lumen and show 4 to 6 nuclei, rather than the 2 to 4 seen in the PCT.
Also, the nuclei of DT cells tend to be closer to the lumen, and to bulge out into it somewhat.
The cells of the DCT have less affinity for cytoplasmic stains (due to smaller organelles contents).
In addition, sections of the DCT are seen much less frequently than sections of the PCT since the DCT is much shorter than the PCT.
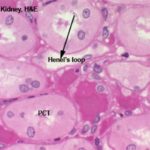
The loop of Henle is made of simple squamous epithelium.
A loop can be identified fairly easily by its thin outer wall and empty lumen.
The loop of Henle could be differentiated from the vasa recta by their regular, rounded shape and the absence of erythrocytes.
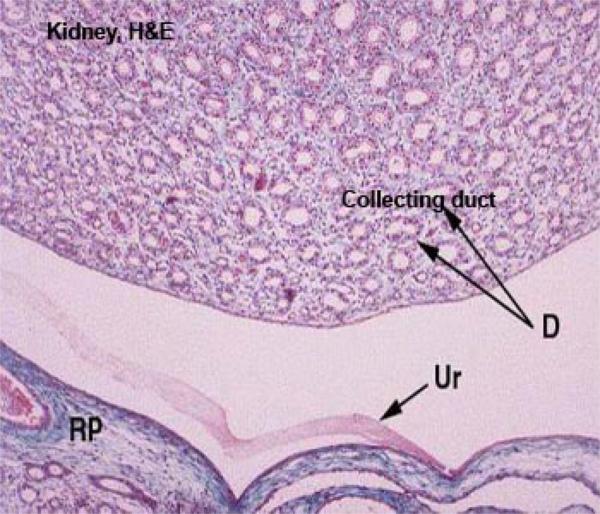
Collecting Tubules and Collecting Ducts ![]()
The collecting tubule receives the filtrate from the DT and carries it down through the medulla and out to the world beyond.
Collecting tubules are quite large, with walls formed of cuboidal epithelium.
They’re larger than the DT and the nuclei of their mural cells bulge somewhat into the lumen. The cells themselves are rather pale staining compared to those of the other tubules.
Large collecting ducts are formed by the fusion of a number of collecting tubules, and drain several nephrons.
Role of the Collecting Ducts
The epithelium of the collecting ducts is sensitive to antidiuretic hormone (ADH). When ADH levels are high, the walls become very permeable to water. When ADH levels are low, the opposite occurs.
In the presence of anti-diuretic hormone (ADH) secreted by the pituitary, the collecting ducts become permeable to water.
Water is drawn out of the collecting tubules, by osmosis, into extracellular fluid and thence into adjacent vasa recta.
Release of ADH is stimulated by dehydration.
This promotes the uptake of water from the collecting tubules thereby producing a reduced volume of hypertonic urine.
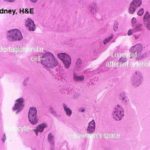
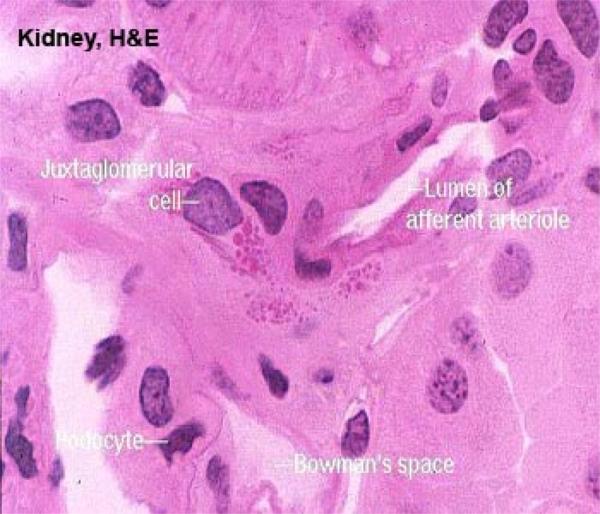
The juxtaglomerular apparatus is involved in the regulation of systemic blood pressure, and hence indirectly controls glomerular blood flow.
In the wall of the DT at the point where the tubule and the afferent arteriole are in contact cells are taller than the normal and much more crowded together. This is the Macula Densa.
The macula densa is only one part of the juxtaglomerular apparatus.
In the wall of the arteriole itself, specialized juxtaglomerular cells form a cuff or sleeve around the lumen.
These cells are modified smooth muscle cells, recognized by the presence of bright pink granules in their cytoplasm.
These are renin granules. Renin is a hormone involved in regulation of blood pressure.
he juxtaglomerular cells in the afferent arteriole are thought to be specialized blood pressure receptors, with the capacity to release renin when needed.
The third component of the juxtaglomerular apparatus is the Lacis cells (or Polkissen) this is small group of cells lies between the macula densa and Bowman’s capsule at the point of entery of the afferent arteriole.
These are thought to be extra-mesangial cells. Lacis cells may produce the hormone erythropoietin which promotes erythropoiesis in bone marrow.
The stimulus for erythropoietin release is postulated to be a low concentration of oxygen in the blood of the afferent arteriole.
A fall in systemic blood pressure triggers a response in the juxtaglomerular cells, which causes renin release.
The macula densa cells may function as chemoreceptors, sensing a drop in sodium concentration of the DT, to trigger renin release by an alternative pathway.
The enzyme renin, when liberated into the bloodstream, acts on the plasma globulin, angiotensinogen to produce angiotensin I.
A further enzyme, called converting enzyme convert the angiotensin I into angiotensin II which is a potent vasoconstrictor.
Angiotensin II brings about an increased blood pressure in two ways: first, by constriction of peripheral blood vessels, and secondly by promoting the release of aldosterone from the adrenal cortex.
Aldosteron increases blood pressure by promoting the reabsorption of sodium ions and therefore water from the DCT, thus expanding the plasma volume and hence increasing blood pressure.
The kidney is a place in which blood is filtered.
The blood flow is the driving force behind urine formation.
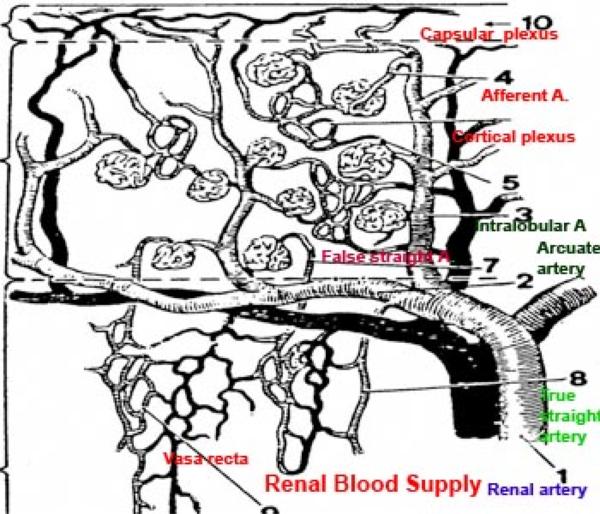
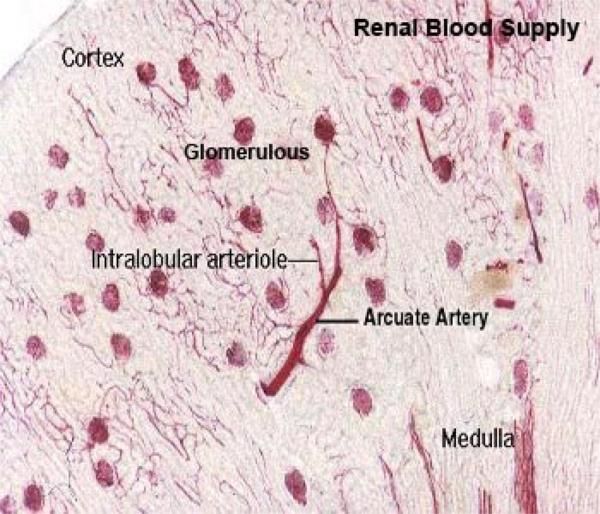
The kidnye is supplied bye renal artery, and is distributed through a series of smaller vessels.
After entering the kidney proper, each renal artery breaks up into several interlobar arteries which course within the renal column between the pyramids.
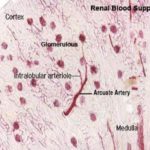
At the cortico-medullary junction, these arteries form branches that arch over the pyramids as the arcuate arteries (so called because they run along the “arc” of the kidney’s curvature) give off smaller branches that supply the peripheral cortex.
Small intralobular arteries are visible in the cortex, running radially outside the level of the arcuate arteries.
From these arise the afferent arterioles which directly supply blood to the capillaries in the renal corpuscles.
These capillary beds (the glomeruli) can be seen “dangling” from afferent arterioles like small berries.
Outflow from the glomeruli is via the efferent arterioles, which are somewhat smaller than the afferent ones.
These almost immediately break up into the fine capillary networks which surround the loops of the nephrons.
The capillaries are vital to maintaining the gradient of sodium concentration in the kidney’s interstitial spaces; the concentration is highest in the deep medulla and lowest in the cortex.
In some places the capillaries are arranged in a net-like configuration, close to the glomeruli from which they arise; and others where capillary beds are arranged as long straight segments. Within these straight regions are recurrent loops.
By appropriate opening and closure of venous shunts the kidney can regulate sodium concentrations by regulating the flow of blood in the capillaries that surround the nephrons proper (the rete mirabile) or their extension, the straight and recurrent vessels of the vasa recta.
The medullary blood supply is derived from the efferent arterioles originating from renal corpuscles close to the medulla.
The vessels extending into the medulla from these efferent arterioles are the false straight arteries as well as vessels originating from the arcuate arteries to the medulla which are called the true straight arteries.
Eventually drainage from all of the post-glomerular capillaries is collected into larger and larger veins until it leave the kidney via the renal vein, at the hilus.
The renal pelvis is the upper end of the ureter, attached just below the tips of the kidney’s lobes.
It catches the urine and drains it out. It’s a CT “cup” lined with urinary epithelium.
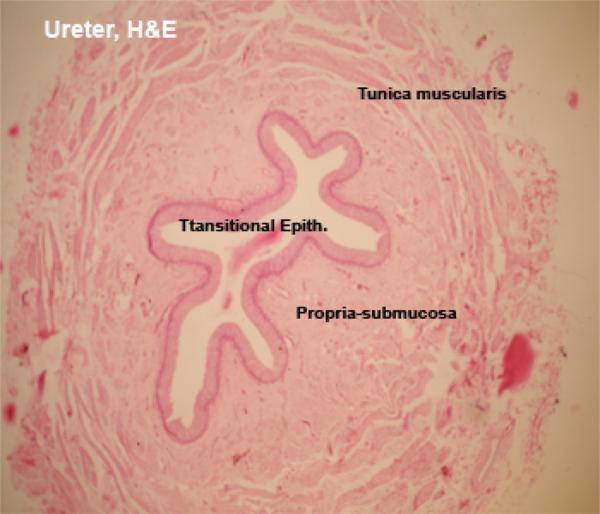
The wall of the ureter is formed of mucosa-submucosa, tunica muscularis and adventitia or serosa.
The mucosa is folded and is lined with transitional epithelium that rests on fibro-elastic connective tissue propria-submucosa which in horse, donkey and mule contains simple branched tubuloalveolar seromucoid glands, the secretion of these glands impart a characteristic turbid appearance for the urine of these species.
The tunica muscularis is formed of three ill-defined layers; inner and outer longitudinal and middle circular layers of smooth muscle fibers separated by loose connective tissue.
Near the bladder, the middle layer disappears
The outer coat of the ureter may be either adventitia or serosa depending on the level of the sections.
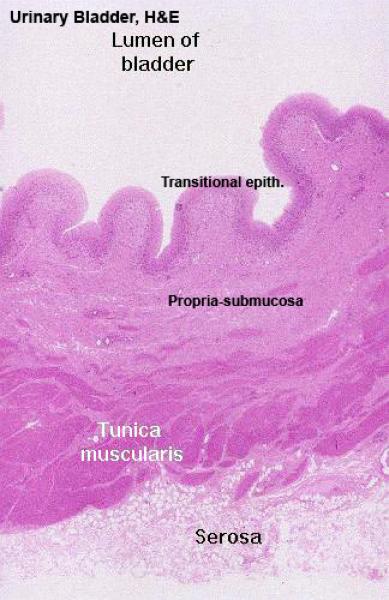
The bladder is a temporary storage organ for urine, whose capacity (in humans) can be measured in terms of milliliters, or in miles between rest stops on the interstate highways.
The wall of the bladder is similar to that of the ureter, but there are said to be three layers of smooth muscle: inner and outer layers are longitudinal, and the middle more or less circular.
There is also a fair amount of elastic connective tissue in the wall to provide for elasticity and distensibility.
The lining is urinary epithelium, extensively folded and puckered in the relaxed state.
The mucosa of the urinary bladder is lined with transitional epithelium.
The lamina propria-submucosa is loose fibro-elastic connective tissue layer.
The muscularis mucosa is absent except ox..
The tunica muscularis is formed of interwoven muscle bundles.
Three-layered muscular coat is found near the bladder neck.
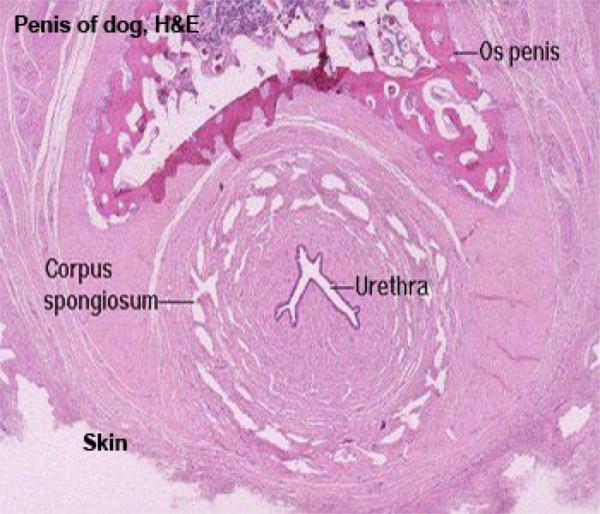
The final passageway, the urethra, which in males does dual duty as part of the urinary tract and the reproductive system.
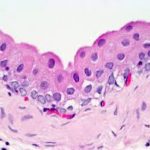
The end of penile urethra has a lining of stratified squamous epithelium; farther up in the pelvic urethra, nearer the bladder, the lining is still urinary epithelium.
The female urethra is the part of the urinary passage and conducts the urine from the urinary bladder to the exterior.
The urethral epithelium begins with transitional type that changes to stratified columnar and ends by non-keratinized stratified squamous epithelium.
The urethral epithelium may show intraepithelial mucous secreting cells and subject to species variations.
The fibro-elastic propria-submucosa is permeated by cavernous spaces with species differences.
The tunica muscularis is represented by bundles of smooth muscles and striated fibers that form inner circular and outer longitudinal layers in the proximal part of the urethra.