-
- Figure 1
-
- Figure 2
-
- Figure 3
-
- Figure 4
-
- Figure 5
-
- Figure 6
-
- Figure 7
-
- Figure 8
-
- Figure 9
-
- Figure 10
-
- Figure 11
-
- Figure 12
-
- Figure 13
-
- Figure 14
-
- Figure 15
-
- Figure 16
-
- Figure 17
-
- Figure 18
-
- Figure 19
-
- Figure 20
Endocrine System
The endocrine system is formed of endocrine glands whose products are secreted into the blood, to affect tissues distant from the site of secretion.
These secretory products are hormones. Because endocrine organs secrete directly into the blood they haven’t got the elaborate duct system exocrine glands have.
Histologically they tend to be quite uniform in structure: masses of cells with blood vessels running through them. Most endocrine organs are “epithelial” and would of course therefore be categorized as glandular epithelium.
Be aware, however, that not every organ that could be considered “endocrine” is necessarily “epithelial.” The brain is quite rightly considered “endocrine” but it’s not an epithelial structure.
The endocrine glands are supported by a little stromal elements.
The endocrine glands exist in the form of discrete endocrine organs, e.g. the pituitary gland, the pineal body, the adrenal glands, and the thyroid and parathyroid glands.
Others endocrine tissues are found in association with exocrine glands, e.g. pancreas or within organs e.g., the kidney, the testis, the ovary, the placenta, the brain and gastrointestinal tract.
In brief, the structure of all endocrine glands conforms to a general pattern:
1. They are composed of secretory cells that are arranged either in the form of cords, clumps or sometimes forms follicles.
2. They are richly vascular always containing dense network of sinusoidal capillaries.
3. They have very little stromal (CT) elements.
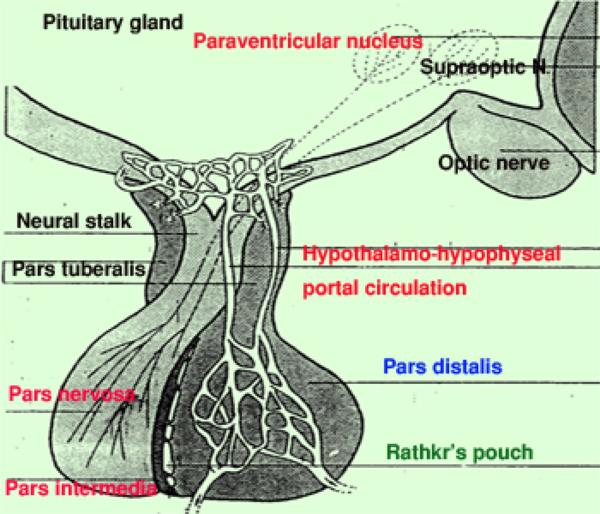
The pituitary gland is a small endocrine gland located beneath the hypothalamus to which it remains attached by pituitary stalk or neural stalk.
The pituitary gland widely referred to as the “Master gland” because its hormonal secretions have as their target organs other endocrine glands.
The gland is almost completely enclosed in a bony cavity in the sphenoid bone called the sella turcica.
The pituitary gland arises from two different embryonic rudiments: one is a depression in the floor of the forming brain and the other is an evagination of the dorsal surface of the forming gut.
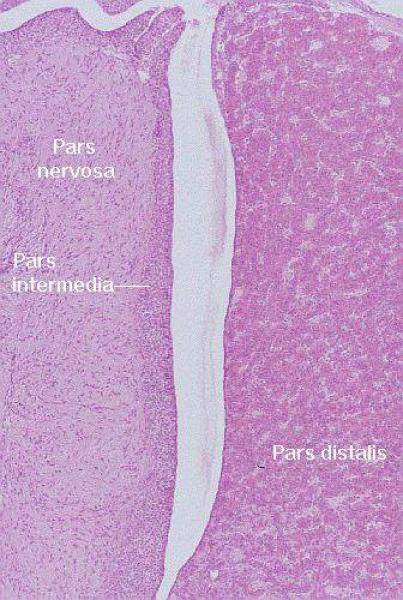
Consequently, in the finished state, this organ has two parts that are distinctly different in appearance and in nature.
Regions of the Pituitary:
The pituitary glands are composed of the adenohypophysis formerly called anterior pituitary and the neurohypophysis formerly called posterior pituitary.
The terms adenohypophysis and neurohypophysis are more accurately reflect the origins of this organ from, respectively, the gut and the neural tube.
Adenohypophysis
The adenohypohysis is that portion of the gland that comes from the gut: it is epithelial in nature.
The root word “adeno” implies its origin from epithelium, not neural tissue.
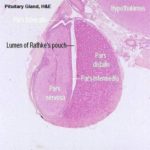
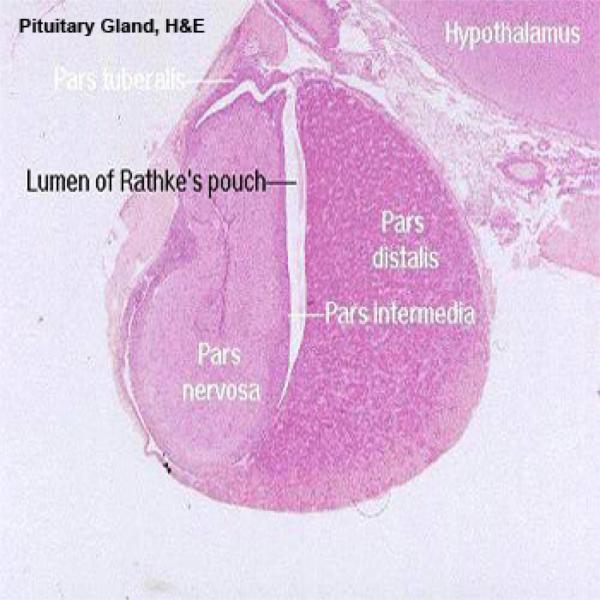
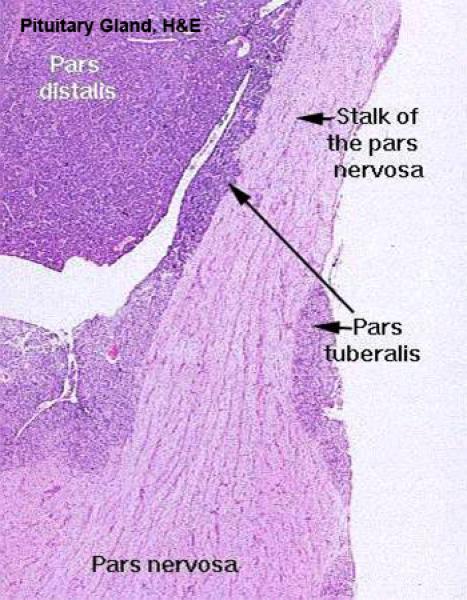
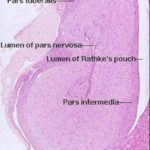
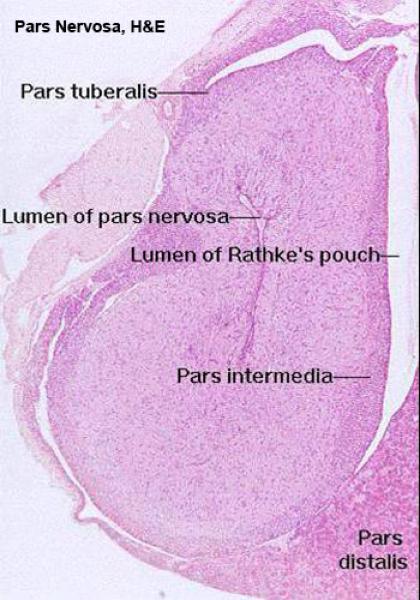
![]() The mature adenohypophysis has several distinct parts: the pars distalis, the pars intermedia, which is physically associated with the neural rudiment, and the pars tuberalis, which comes to form a sort of sleeve-like covering of the stalk from which the neural portion is dependent.
The mature adenohypophysis has several distinct parts: the pars distalis, the pars intermedia, which is physically associated with the neural rudiment, and the pars tuberalis, which comes to form a sort of sleeve-like covering of the stalk from which the neural portion is dependent.
In some species, there may be visible an opening or gap between the pars distalis and the pars intermedia.
This–“Rathke’s Pouch”–is the vestigial remnant of the lumen of the gut pouch. It has no functional significance and in some species it’s obliterated.
Neurohypophysis
The neurohypophysis is that part of the organ derived from the brain, and unlike the adenohypophysis, it retains its connection to the tissue of origin, via long stalk.
The neurohypophysis has a lumen, and that lumen is continuous with the lumen of the brain’s third ventricle.
The lumen of the neurohypophysis, like the ventricles of the brain, is lined by ependymal cells.
This part of the pituitary gland, in an anatomic sense, may be considered a part of the brain, and its histologic appearance confirms this.
It consists mainly of nerve fibers with associated glial elements.
It is a dependent part of the median eminence of the hypothalamus.
The cells of each pituitary region have distinct hormonal products associated with them.
Microscopic Anatomy of the Pituitary
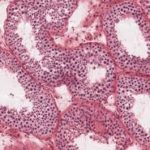
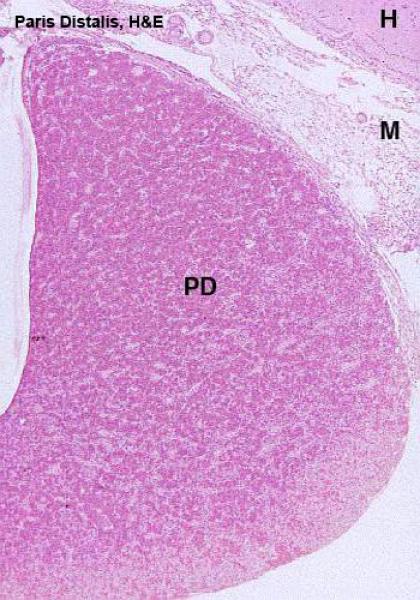
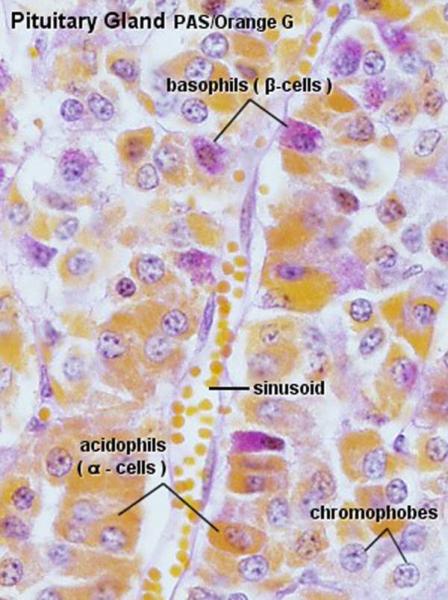
The pars distalis is formed of secretory cells arranged into irregular cords or clusters separated by a rich network of fenestrated sinusoidal capillaries.
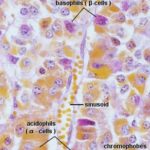
The cells are classified into two groups, chromophils and chromophobes, according to their staining affinities for histological dyes.
The chromophils are cells with variable size with strong affinity for histological dyes whereas the chromophobes are smaller cells which have little affinity for stains.
Chromophils ![]()
Traditionally, the chromophils have been subdivided into two groups: acidophils and basophils because of their staining properties with a variety of histological methods.
In H & E sections, the distinction between acidophils and basophils is much less obvious.
Using EM and immunohistochemical techniques, five cell types are recognized according to their secretory products: somatotrophs, mammotrophs, corticotrophs, thyrotrophs and gonadotrophs.
1.Somatotrophs ![]()
They are the most numerous types, making up half of the cells forming the pars distalis.
They are acidophilic at LM level and have cytoplasmic granules that stain orange with orange-G stain.
With EM, they are spherical or ovoid cells with fairly well developed rough-surfaced endoplasmic reticulum and abundant relatively large secretory granules that are approximately 350 nm in diameter.
The somatotrophs secrete growth hormone which is essential for adequate growth of chondrocytes and adequate secretion of cartilage matrix in the epiphyseal plates of long bones.
Defective growth due to growth hormone deficiency is often referred to as pituitary dwarfism.
Excessive growth may occur due to continuous unregulated production of growth hormone (somatotrophs tumor) after adolescence and is referred to pituitary gigantism.
2. Mammotrophs ![]()
They comprise up to 20 % of the cells forming the pars distalis.
hey are acidophilic at the LM level and have cytoplasmic granules that stain red with azocarmine.
With EM, the cells are small, spindle-shaped with only a few fairly small (about 200 nm in diameter) cytoplasmic granules.
They secrete prolactin hormone which stimulate the mammary gland to produce milk. Therefore, the number of these cells as well as the number of their granules tends to increase during pregnancy and lactation.
This hormone is also associated with maternal behaviors such as broodness in chickens and production of crop milk in pigeons and doves.
3. Corticotrophs ![]()
They constitute about 20% of the pars distalis.
They are basophilic at the LM level.
With EM, the cells are typically spherical to ovoid, contain granules that are only approximately 100 nm to 200 nm in diameter and that show noticeable variation both in their electron density and in their size.
The chief hormone produced by corticotrophs is adrenocorticotrophic hormone (ACTH).
4. Thyrotrophs ![]()
They constitute only 5 % of the pars distalis mass.
They are basophilic at the LM level, their cytoplasmic granules are stainable with aldehyde fuchsin and alcian blue.
With EM, the cells have considerably smaller, peripherally located granules.
They produce the glycoprotein hormone thyroid stimulating hormone (TSH) which exerts a trophic influence on thyroid follicular cells.
5. Gonadotrophs ![]()
They make about 5% of the pars distalis mass.
They are basophilic at the LM level and their cytoplasmic granules are stainable with PAS and alcian blue.
With EM, the cells are fusiform with eccentric nuclei and heterogenous contents of secretory granules with a medium size range (200-400 nm) and variable electron density.
They secrete gonadotrophins (FSH and LH). The functions of gonadotrophins are to stimulate the gonads so as to bring about the development and maturation of germ cells and the secretion of sex hormones.
Pars Intermedia and Pars Tuberalis ![]()
The pars intermedia is pressed up close to the neurohypophysis.
It too is an epithelial area, but its staining reaction is distinctly different from the pars distalis.
There are normally no acidophilic cells obvious in it.
The arrangement of the cords and sinuses is somewhat different too, and you should have no trouble telling this region from the pars distalis. However, the pars intermedia grades into the pars tuberalis without a definite boundary.
The pars intermedia secretes MSH, or “intermedin,” which has some similarities to ACTH in structure and in activity.
The pars intermedia is more extensive in the domestic animals than in humans.
The cleft next to the pars intermedia is the remnant of the embryonic Rathke’s pouch, the evagination of the gut tube that forms one rudiment of the organ.
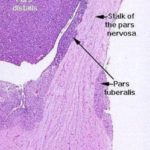
The pars tuberalis is not yet known to secrete any hormones.
Even though it may be similar to the pars intermedia, the pars tuberalis is quite clearly distinct from the adjacent pars nervosa; it forms a “cuff” or “sleeve” around the stalk of the pars nervosa itself.
The pars nervosa arises as a ventral outpouching of the floor of the brain, and it retains its connection with the brain.
It has a long, narrow lumen in it. The lining cells of the pars nervosa are the same sort of ependymal cells found lining the central nervous system.
If you have a favorable section, you may be able to trace the connection between the space in the pars nervosa and the brain ventricle.
The pars nervosa is neural tissue.
It consists of well vascularized fiber tracts. Most of these are axons coming from neurons whose cell bodies are located in the hypothalamus.
These axons carry materials made in the brain down into the deep part of the pars nervosa.
Unlike most other axons, they don’t form synapses with other neurons, however.
When these axons reach the pars nervosa they end in contact with blood capillaries.
The axons are supported by neuroglial cells called pituicytes.
In H & E sections, the axons are indistinguishable from the cytoplasm of the pituicytes.
Enlargement of neurosecretory axons are referred to as Herring bodies which are sites of storage and disposal of hormones.
The pars nervosa is a place of storage and release of hormones made in the brain.
The two hormones associated with it are oxytocin and vasopressin. Instead of neurotransmitters, these hormones are made in the brain’s supraoptic and paraventricular areas, then carried to the pars nervosa in the axons that end at the surface of capillaries.
Upon appropriate stimulation, they release their products directly into the blood at these sites.
Thus the brain is acting as an endocrine organ, and releasing its products at a dependent site.
Vascular Considerations and Their Relationship to Pituitary Function
The entire hypophysis is extensively vascularized.
The adenohypophysis in particular, there are irregularly shaped sinusoids between the cords of cells in the pars intermedia, the pars distalis, and the pars tuberalis.
Sinusoids are essentially capillaries, and the blood flowing through these obviously will have direct contact with the adjacent cells of the cords in the adenohypophysis.
Portal Circulation and the Adenohypophysis ![]()
The pituitary receives blood from branches of the internal carotid artery.
The bulk of this supply goes first to the median eminence of the brain, where it feeds an extensive capillary plexus.
As with any capillary bed, the next step is to form small venules; but the veins in this case don’t simply drain into the right side of the circulatory system directly.
Instead, they drain into the blood sinusoids of the adenohypophysis.
This arrangement–a capillary bed draining into a second capillary bed via a short set of venules–is a portal system, and it’s vital to normal function.
The key point here is that anything put into the “upstream” end of the portal system in the neurohypophysis is immediately transported to the adenohypophysis.
The neurons of the brain, particularly those of the hypothalamus are secretory cells.
Some of them secrete not neurotransmitters, but hormones, and these hormones have as their target organ the cells of the adenohypophysis.
These neuronal releasing factors are chemical signals that trigger the release of other hormones from cells in the non-neural portions of the gland.
The hormones of the pars distalis and intermedia are released into the blood in response to the releasing factors.
These hormones in turn target other organs, including other endocrine organs.
It should be obvious from the above discussion that the pituitary is not in any sense a “Master Gland”.
The brain (through the agency of the pituitary gland) is what really controls the output of the endocrine system.
In the case of oxytocin and vasopressin, the brain actually bypasses the pituitary entirely and dumps hormones directly into the blood.
The pituitary is, at best, a sort of “Overseer Gland” whose job it is to transmit the orders of the true “Master Gland,” the brain, to the rest of the organs.
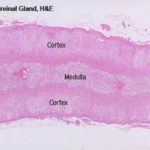
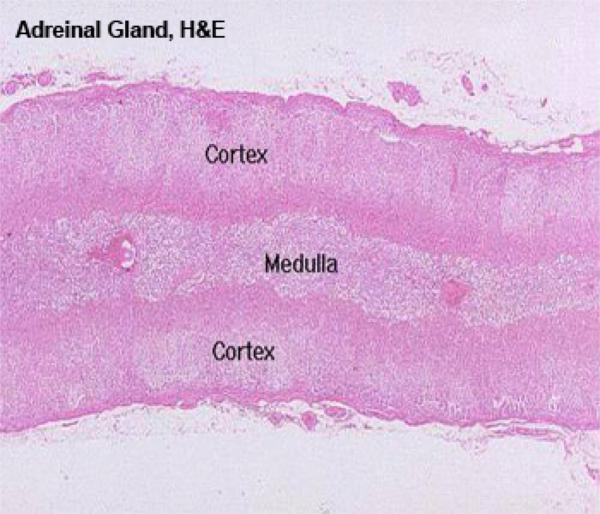
The adrenal gland has a distinct cortex and medulla, each of different embryologic origin and different function.
Blood Supply
The adrenal cortex and the medulla have separate and distinct arterial supplies, and this is related to the different origins and functions of the two parts.
The medullary arteries run straight through the cortex, bypassing it completely, and ramify into the sinusoids of the medulla.
The cortex is supplied by cortical arteries which enter the organ via the capsule and discharge their blood into the sinusoids of the zona glomerulosa.
Venous outflow from the cortex is mixed with that of the medulla, and carries secretions from both parts.
The Adrenal Cortex
The adrenal gland as a whole is surrounded by a distinct CT capsule, which sends a few septa into the parenchyma of this gland; generally, though, there is little internal lobulation.
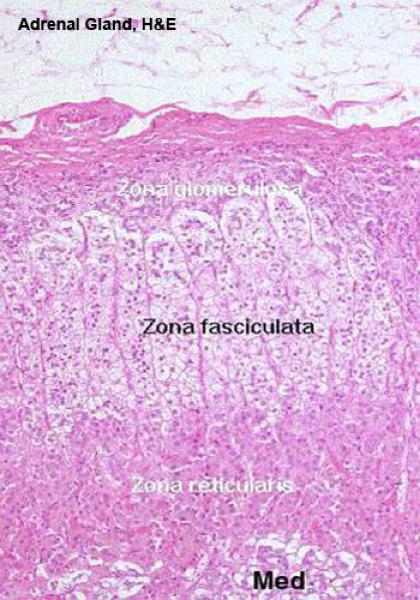
The layers of cells immediately beneath the capsule are organized into the zona glomerulosa, or “region of little globes.”
In ruminants, the secretory cells are arranged in irregular ovoid clusters and cords separated by delicate connective tissue septa containing capillary sinusoids.
In solipeds (horses, donkeys and mules), carnivores and pigs, the cells are arranged in arched clusters, hence the name zona arcuata.
The secretory cells have spherical darkly stained nuclei and acidophilic cytoplasm.
With EM, The cytoplasm contains much sER, numerous mitochondria and lipid droplets) steroid-secreting cells.(
The cells of zona glomerulosa secrete the mineralocorticoids hormone (aldosterone) which regulates sodium and potassium ions levels as well as participates in blood pressure regulation.
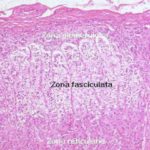
Zona Fasciculata ![]()
It is the intermediate and broadest of the three zones of the adrenal cortex.
It consists of parallel cords of secretory cells arranged at right angles to the capsule.
The cords are often only one cell thick and separated by connective tissue strands containing capillary sinusoids.
The secretory cells are polyhedral with central lightly stained nuclei. The cytoplasm is rich in sER and lipid droplets, therefore it appears vacuolated and so they are called spongiocytes.
The cells of the zona fasiculata secrete glucocorticoids which regulate the carbohydrate metabolism.
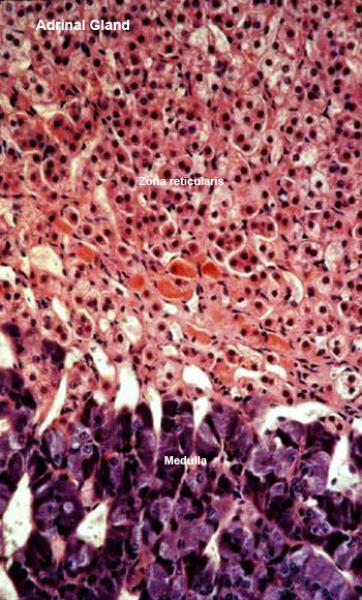
Zona Reticularis ![]()
The cells in that region of the cortex closest to the medulla lose this regular arrangement, and are organized into anastomosing cords to form a network-like zona reticularis.
Separating the rows and cords of cells are irregular vascular channels, or sinusoids similar to those in the liver.
The cells of the zona reticularis secrete small quantities of steroid sex hormones.
The adrenal medulla comes from an entirely different embryonic rudiment than does the cortex.
The cortex arises from a condensation of embryonic mesoderm, but the medulla, ultimately, is of neuroectodermal origin.
As a result it has different properties and its secretions are triggered by different stimuli.
The cells of the adrenal medulla are somewhat more basophilic than those of the cortex. With appropriate stains, these medullary cells show the so called “chromaffin reaction” characteristic of cells producing catecholamine compounds, which of course is what they are doing.
The adrenal medulla makes the catecholamine hormones epinephrine and norepinephrine.
The adrenal medulla comes from an aggregation of neural crest cells. In the fully formed organ, innervation of the medulla by the autonomic nervous system is direct; there is no intervening ANS ganglion as in other target organs.
In reality, the adrenal medulla can be though of as a sort of ganglion combined with its effector organ.
Functional Implications of Adrenal Gland Structure
The adrenal gland’s dual blood supply has significant consequences.
The cortex, like any ordinary endocrine gland, responds to a pituitary hormone than directs the cells to put their product into the blood stream.
The pituitary hormone ACTH is brought to the cortex via the cortical arteries, passes into the sinusoids, and stimulates release of steroid hormones from the cortical cells back into the sinusoids and out to the rest of the body.
The medulla however, does not need and does not have any such chemical messenger.
It is stimulated by direct neural signals. Axons from the autonomic nervous system directly synapse with cells of the medulla, and if those axons discharge, the cells respond by releasing their products into the blood flowing through the sinusoids in the medulla.
The most familiar example of medullary action is the “flight or fight” reaction.
Imagine you are walking, and a large bear pops out from behind a bush 10 feet from you.
Your eyes perceive the image of the bear; your brain integrates the information and decodes the image BEAR!
Immediately your sympathetic nervous system, in response to the information that a B-B-B-BEAR!! is standing in front of you sends a message to the adrenal medulla; the adrenal medulla releases epinephrine into the blood.
This hormone–also called “adrenaline“–has a number of target organs.
It causes your gut musculature to stop peristalsis; your pupils to dilate; your circulatory system to start sending plenty of oxygenated blood to your limbs.
Thus, when you turn and run as fast as you can away from that B-B-B-B-B-BEAR!!! you are putting maximum effort into the escape.(
Epinephrine receptors are present on nearly all cells, and the adaptation of this material which is in some places known to be a neurotransmitter) to this use makes it a sort of “general quarters alarm” for the entire organism.
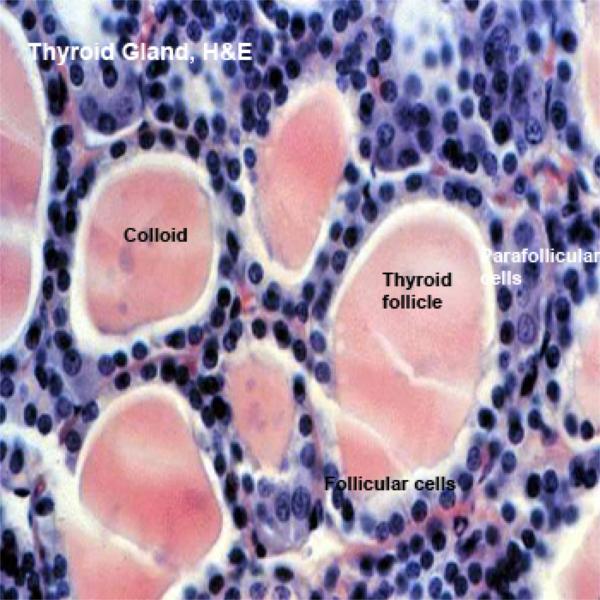
The thyroid is a highly vascular bilobed gland located in the neck in front of the upper part of the trachea.
The gland is enveloped by a loose connective tissue capsule sending septa interiorly dividing the gland into lobules, and carry a rich blood supply together with lymphatics and nerves.
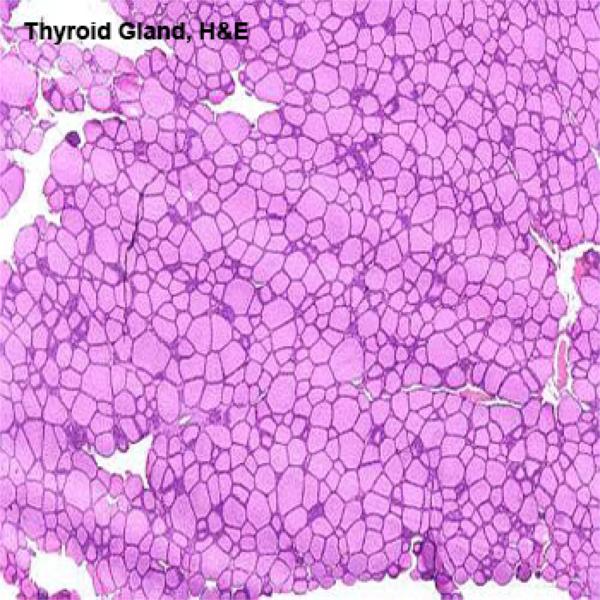
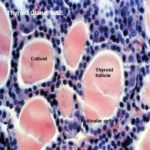
Each lobule is composed of a large number of irregular spheroidal structures known as thyroid follicles which are packed closely together in a delicate reticular network containing many capillaries.
The wall of the follicles is composed of simple cuboidal epithelium.
The amorphous material present inside the follicles is colloid, an inactive storage form of the gland’s secretion.
The thyroid is unique among endocrine glands in that it stores its secretory product extracellularly. Indeed, it’s unusual for an endocrine gland to store products at all.
Thyroid Follicular Cells ![]()
With EM, the cells have apical microvilli on their lumina surface.
They exhibit obvious signs of active glycoprotein secreting cells.
This include the presence of widely dilated cisternae of rER and distended Golgi saccules and the accumulation of apical secretory granules that discharge their glycoprotein content by exocytosis into the follicular lumen.
Besides producing thyroglobulin, these cells are able to trap iodide from the bloodstream by means of active transport mechanism.
Furthermore, they produce a peroxidase that is uses to oxidize this iodide.
The oxidized iodine is utilized to iodinate the tyrosyl groups of the noniodinated thyroglobulin.
Iodination occurs intraluminally.
Under the influence of TSH, the follicular cells actively take up iodinated thyroglobulin by endocytosis from the lumen of each thyroid follicle and then submit it to lysosomal digestion.
Two products liberated as a result of intracellular proteolysis are T3 and T4.
They diffuse out of the cells, where they enter the circulation by way of the numerous fenestrated capillaries that are closely associated with theses cells.
In active thyroid gland, the follicles tend to be small, the cells are tall cuboidal and the amount of colloid decreases.
In less active gland, the follicles are distended with colloid and the lining cells appear flattened.
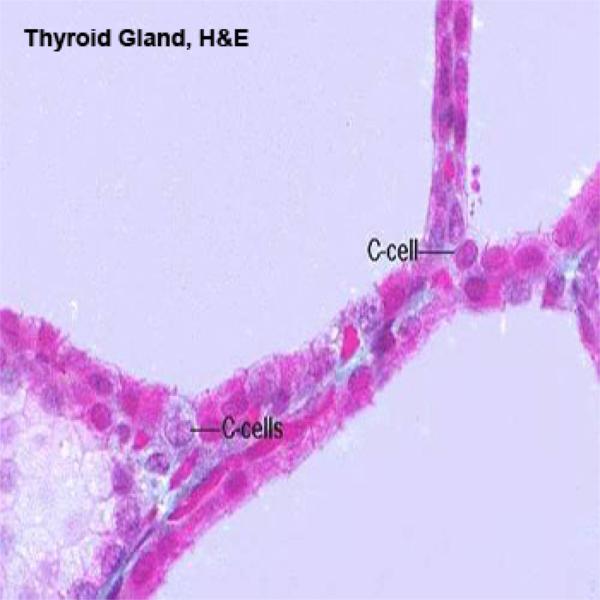
Thyroid Parafollicula Cells or C Cells ![]()
They are found either as single cell located among the follicular cells or as small clusters in the interfollicular spaces.
They are large, rounded pale-staining cells that have a central spherical nucleus.
With EM, these cells lie enclosed within the same basement membrane as the follicular epithelia cells.
Their cytoplasm contains abundant spherical secretory granules 100-200 nm in diameter, that contain the polypeptide hormone calcitonin.
Calcitonin is a blood-calcium lowering hormone that has an antagonistic action to parathyroid hormone.
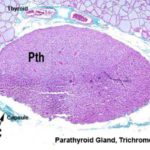
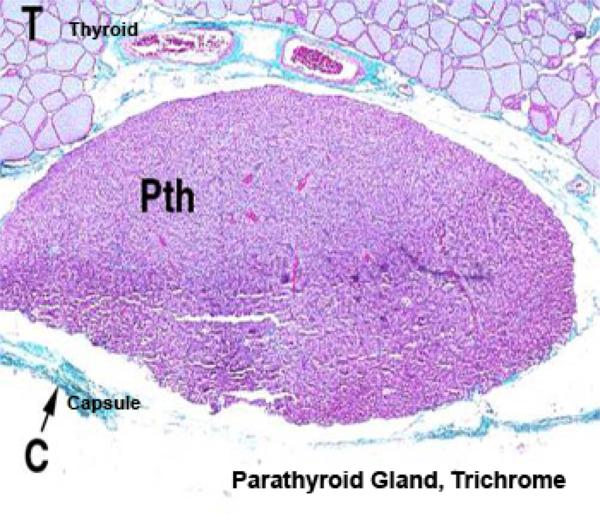
The parathyroid gland are four small oval bodies associated with the thyroid gland, but which are separate endocrine organs in and of themselves.
It has its own distinct capsule, and is not lobulated as is the thyroid gland.
Like other endocrine organs it’s well vascularized.
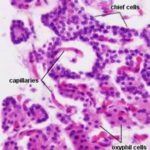
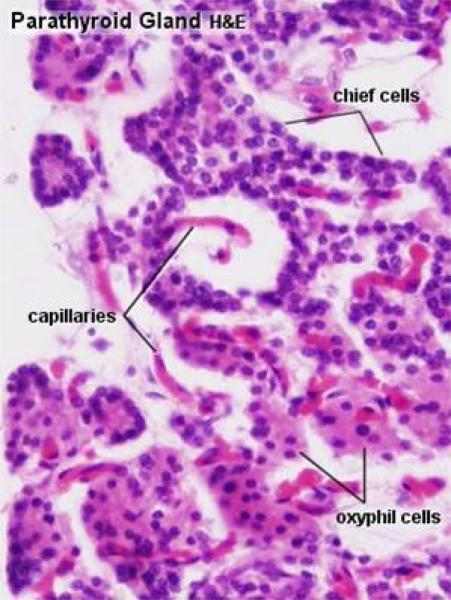
The cells forming the parathyroid gland are of two main types: chief cells and oxphil cells.
1. Chief cells ![]()
They are the most numerous cell type.
The cells are small polygonal in shape with prominent nuclei and relatively little cytoplasm.
Active cells contain much rER and stain strongly.
Less active cells contain little rER and stain poorly.
The chief cells produce parathyroid hormone (or parathormone, as it’s sometimes called, abbreviated PTH) which is a blood calcium-raising hormone.
2. Oxphil cells ![]()
They are larger and much less numerous than chief cells.
They tend to occur in clumps.
he cells have smaller, darkly stained nuclei and strongly acidophilic cytoplasm.
They do not secrete hormone, their functions are poorly understood.
Functional Considerations
The detailed physiology of the thyroid and parathyroid glands are extremely important clinically.
The thyroid gland’s principal product, thyroxin, effects an increase in metabolic rate.
In some animals under prolonged cold stress, such as Arctic wolves and sled dogs, thyroxin output is often elevated to increase the rate of body heat generation. Such dogs will sleep outdoors in weather as cold as -40 C°!
The calcitonin produced by the C-cells brings about a decrease in blood calcium levels, and an increase in the deposition of calcium in bone.
This effect is antagonized by parathyroid hormone, which acts to release calcium from bone and increase its concentration in the blood.
The pineal gland sometimes called the epiphysis cerebri is a sort of cone shaped structure in the midline of the brain, projecting from the roof of the diencephalon.
The name comes from the Latin pinus, a pine tree, from its vague resemblance to one of these.
Histologically speaking, the pineal gland is uninteresting; about 95% of the cells are the pinealocytes, the functional cells which manufacture the pineal hormone melatonin (5-methoxy-N-acetyl-tryptamine).
These cells are large and lightly stained in H&E, with a spherical nucleus.
The remaining cells of the organ (about 5%) are astrocytes, which cannot be seen on H&E stained slide.
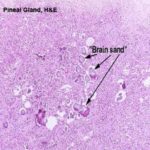
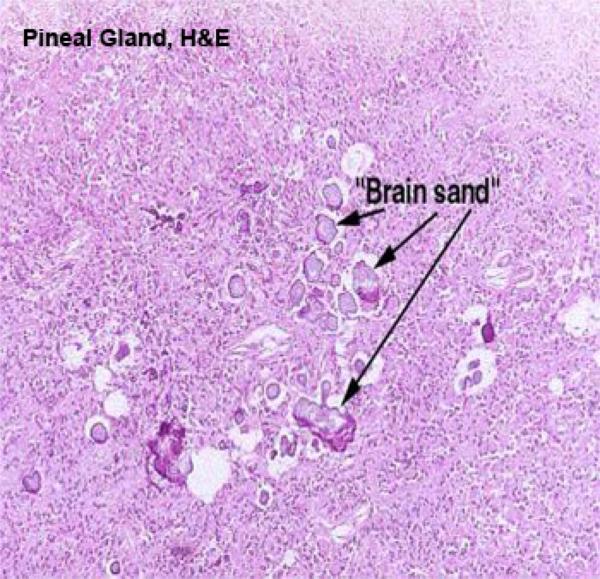
The most obvious aspect of the histology of the pineal gland is the presence of the acervuli cerebri (from Latin, “acervus,” a heap).
This is the so called “brain sand”, and in reality it consists of calcareous secretions that are easily seen with the microscope.
These are not a degenerative change; they’re present in almost all adult mammals and are perfectly normal.
Their mineral composition makes them radio opaque and they have some clinical significance.
The adult pineal is a radiological marker for the midline of the brain. Displacement is easily seen on radiographs, and is a possible indication of the growth of a tumor.
The melatonin, which is an active derivative of serotonin with the capacity to exert a supressive effect on gonadotrophin secretion.
Pineal secretion has the effect of retarding gonadal growth and function.
Melatonin is produced in the pineal only during period when no light entering the eyes.
Endocrine Tissues of the Reproductive System
Both the testis and the ovary are important endocrine organs, and the hormones they produce are important in normal sexual activity, in maturation of the adult body form, and in controlling estrus and pregnancy.
The pancreas in light of its exocrine function is the source of digestive enzymes.
But this organ is also an endocrine gland.
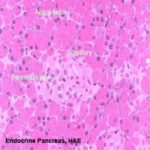
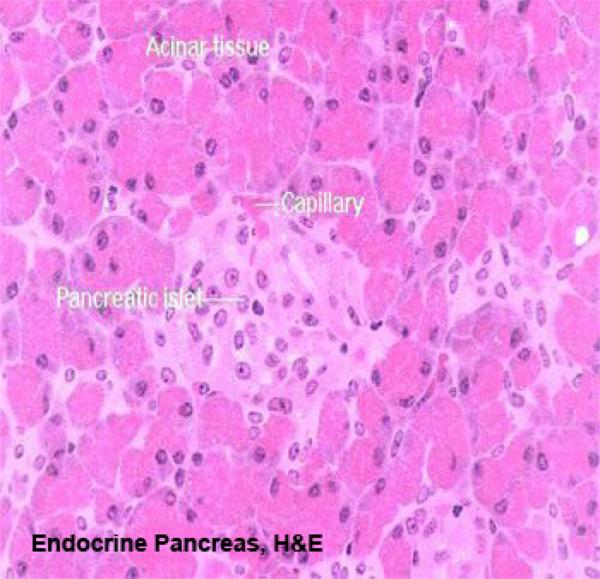
While the bulk of the pancreatic tissue is exocrine in nature, the endocrine pancreas is distributed through the mass of the organ as “islands” of lighter staining material.
These pancreatic islets are easily seen at low power.
At higher magnification several characteristics of the islets can be seen.
Islets are more or less set off from the surrounding exocrine tissue by a thin “capsule” of CT. They’re extensively vascularized, much more so than the surrounding exocrine tissue: an islet is essentially a capillary bed surrounded by secretory cells.
The cells of the islets are not easily distinguished from one another in H&E preparations, but there are six types known to exist.
Alpha Cells (A Cells)
They are large, less numerous making up to 15 % of the islet cell mass.
The cytoplasm contains granules that are insoluble in alcohol and stain brilliant with Masson’s trichrome method.
Generally, the cells tend to be distributed towards the periphery of the islets.
The alpha cells produce glucagon which increases the blood glucose level.
Beta Cell (B Cells)
They are the most numerous cell type and make up to 75% of the total islets cell population) up to 85% in sheep(.
The cells are polyhedral in shape with indistinct cell boundaries.
The cytoplasm contains granules soluble in alcohol and stain dark orange with Mallory’s trichrome stain.
The nuclei are spherical and smaller than that of the alpha cells.
With EM, the beta cell granules are nearly similar to that of the alpha cells.
In dogs, the beta cell granules contain crystalloid structure of variable shape (crystalline insulin) embedded in a pale matrix.
The beta cells secrete insulin which decrease blood glucose level.
Insulin is secreted in response to an elevated blood sugar level. It has two principal actions: 1) to promote glucose uptake in a number of different cell types, and 2) to lower the blood glucose level whenever this level rises.
In diabetes mellitus, there is insulin deficiency leading to elevation of the blood glucose level (i.e., hyperglycemia.
This condition is also characterized by the presence of glucose in the urine (i.e., glycosuria(
Delta Cells (D Cells)
They constitute up 10 % of the islets cell mass.
They are rare in domestic animals.
Compared wit A and B cell, D cells have rather large secretory granules that are less electron dense, and their secretory organelles are less prominent.
The D cells secrete somatostatin which inhibit the release of growth hormone. It also inhibits the release of insulin, glucagon and even somatostatin itself.
C Cells
They are found in guinea pigs and are believed to represent a variety of alpha cells.
F Cells
They occur in the dog’s uncinate process.
Enterochromaffin-like Cells
They secrete serotonin, motilin, and substance P.
Other Endocrine Glands
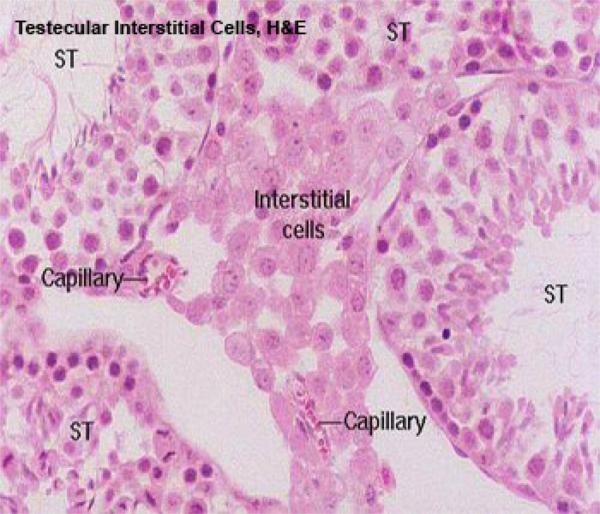
The interstitial cells are small clusters of polygonal cells located in the intercellular spaces between the seminiferous tubules.
They are the source of the male steroid hormone testosterone.
Ultrastructural examination will show that the interstitial cells have the characteristic features associated with steroid synthesis, especially the presence of large quantities of smooth endoplasmic reticulum.
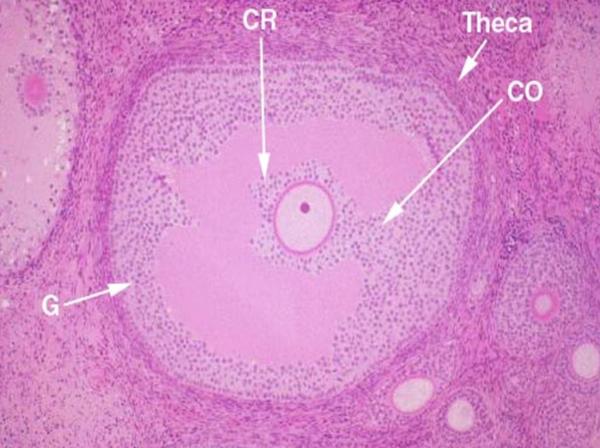
Ovary and Corpus Luteum ![]()
The ovary is a major endocrine organ, and the variations of hormones in the female reproductive cycle are of great importance.They will be considered in details with the female genital system.