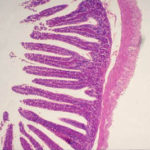
-
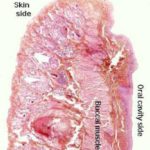
- Figure 1
-
- Figure 2
-
- Figure 3
-
- Figure 4
-
- Figure 5
-
- Figure 6
-
- Figure 7
-
- Figure 8
-
- Figure 9
-
- Figure 10
-
- Figure 11
-
- Figure 12
-
- Figure Figure 13
-
- Figure 14
-
- Figure 15
-
- Figure 16
-
- Figure 17
-
- Figure 18
-
- Figure 19
-
- Figure 20
-
- Figure 21
-
- Figure 22
-
- Figure 23
-
- Figure 24
-
- Figure 25
-
- Figure 26
-
- Figure 27
-
- Figure 28
-
- Figure 29
-
- Figure 30
-
- Figure 31
-
- Figure 32
-
- Figure 33
-
- Figure 34
-
- Figure 35
-
- Figure 36
-
- Figure 37
-
- Figure 38
-
- Figure 39
Digestive System
The digestive system is formed of a digestive tract (gut) and associated glands that include salivary glands, liver and pancreas.
Oral Cavity
The oral cavity is the first portion of the digestive tract, and consists of the lips, teeth, tongue, oral mucosa, and associated salivary glands.
The principal functions of the oral cavity are ingestion, fragmentation and moistening of food. In addition, the oral cavity is involved in speech, facial expression (human), sensory reception and breathing.
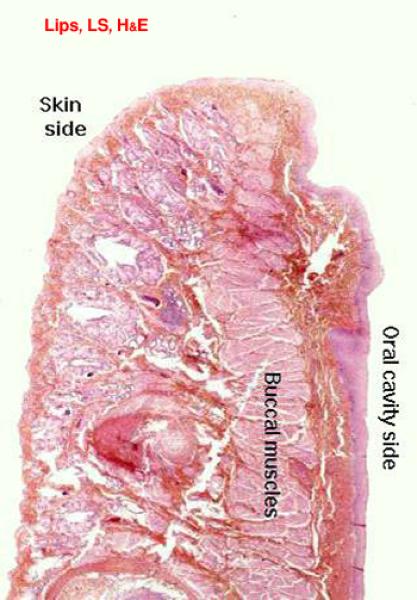
Lips and Cheeks
The external surface (skin side) of the lips and cheeks is covered by hair-bearing skin with sebaceous and sweat glands. ![]()
The internal surface (oral side) is lined by oral mucosa that is made up of stratified squamous epithelium rests on lamina propria-submucosa.
Depending on the nature of the animal’s diet, the epithelium may or may not be keratinized.
Those animals which eat a great deal of roughage, such as ruminants and horses, usually have a very heavily keratinized oral epithelium, while those which eat softer food such as carnivores show less or non keratinized type.
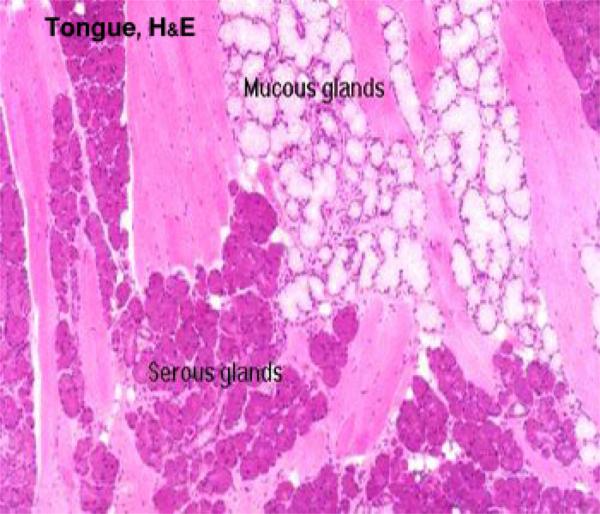
The lamina propria-submucosa is a loose connective tissue layer containing numerous accessory salivary glands of serous, mucous or seromucoid type.
The center of the lips and cheeks contains some of the buccal muscles, strands of skeletal muscle that are part of the muscle sheet underlying the skin.
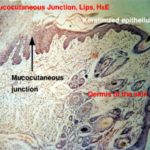
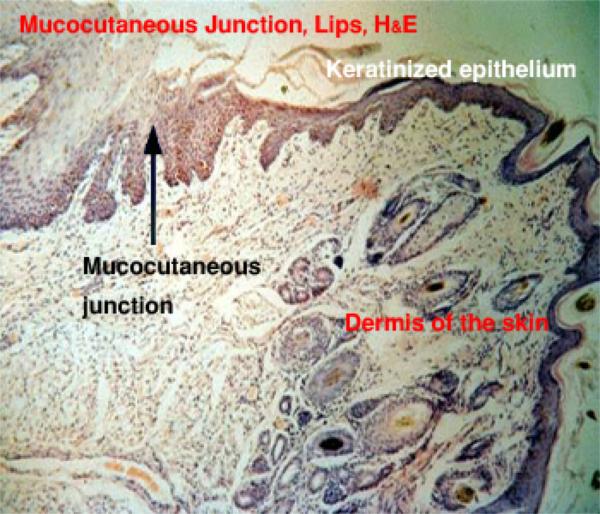
The place of continuity between the “dry” integument and the “wet” oral mucosa is a sort of transition point referred to as a muco-cutaneous junction. ![]()
Tongue
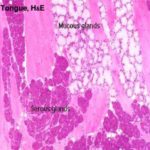
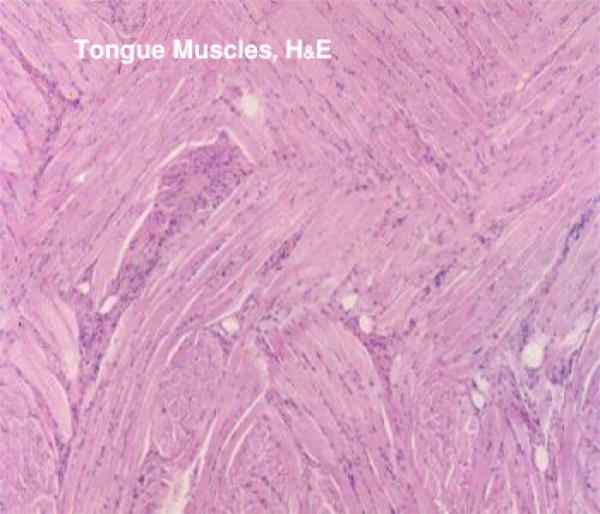
The tongue in mammals is an extremely muscular organ within whose substance there are a number of smaller salivary glands ![]() , and a number of motor and sensory nerve fibers.
, and a number of motor and sensory nerve fibers.
The bulk of the tongue is skeletal muscle, arranged in three layers, all at right angles to each other. This provides for an amazing degree of flexibility and is vital to vocalization. ![]()
There may be a considerable amount of adipose tissue present as well.
The ventral surface of the tongue is smooth and is covered by thin non-keratinized stratified squamous epithelium continuous with that of the floor of the mouth.
The dorsal surface is covered by stratified squamous keratinized epithelium and is raised into a series of elevations called lingual papillae.
Lingual Papilla
Lingual papillae are small elevations found on the dorsal surface of the tongue.
Each papilla is formed of an epithelial cap and a connective tissue core derived from the underlying lamina propria.
They are classified into two major groups: 1. mechanical and 2. gustatory papillae.
The mechanical papillae contain no taste buds and are principally concerned with movement of foods within and into the oral cavity. They include filiform, conical and lenticular papillae.
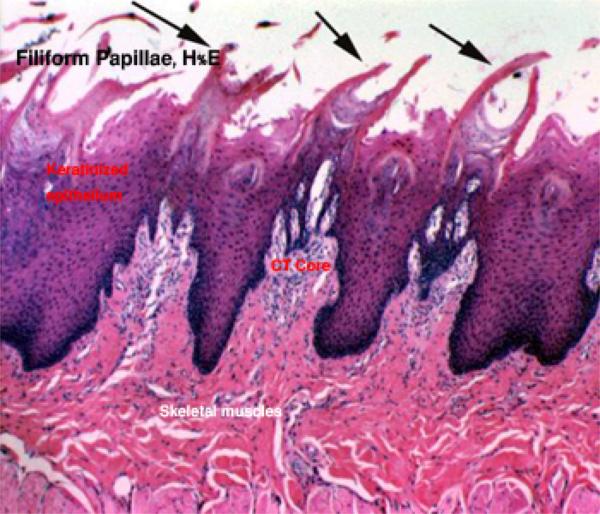
Filiform Papillae
The filiform papillae are the most numerous types, forming a velvety covering on the tongue.
Their shapes vary among different animal species. ![]()
Equine filiform papillae appear as thin cornified threads projecting above the surface and their connective tissue cores end at the bases of the cornified threads.
Ruminant filiform papillae are cone-shaped structure projecting above the surface and their connective tissue cores have several secondary papillae.
Cat filiform papillae are large with two prominences of unequal size. The caudal prominence is especially large and gives rise to a caudally directed keratinized spine. The rostral papilla is rounded with a thinner stratum corneum.
Dog filiform papillae have two or more apices; the caudal one is larger and has a thicker stratum corneum than the others.
Conical Papillae
They are cone-shaped structures, larger than filiform papillae and not highly keratinized.
Their connective tissue cores have both primary and secondary papillae.
They occur on the root of the tongue in dog, cat and pig.
In other species, they occur on the inner side of the cheeks and floor of the oral cavity.
Lenticular papillae
They are flattened lentil-shaped projections occur mainly in ruminants.
- Gustatory Papillae
The gustatory papillae contain taste buds and are primary concerned with reception of taste sensation. They include, vallate, fungiform and foliate papillae.
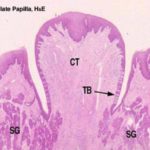
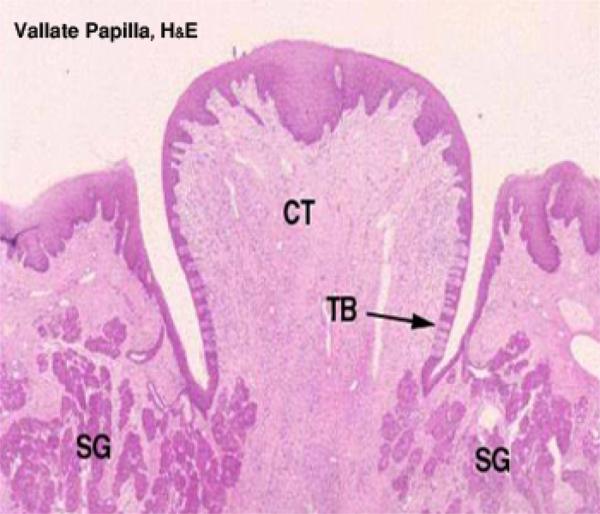
Vallate or Circumvallate Papillae
Vallate papillae are the largest type, easily visible with the naked eye in most animals.
They are paired and located near the back of the tongue. ![]()
A vallate papilla is set into a deep pocket in the tongue’s surface, and anchored at the bottom by a short broad stalk.
It does not protrude above the general level of the surface.
It’s surrounded by a deep narrow epithelial-lined cleft called “moat”.
Vallate papillae usually show taste buds which are located on the undersides of the papilla proper, and on the tongue side of the “moat.”
Aggregates of serous glands called Von-Ebners glands whose ducts open into the moat are located beneath the papilla.
The serous secretions of these associated glands help to clean up the moat and thus facilitate taste reception.
Fungiform Papillae
The fungiform papillae are scattered randomly among the filiform papillae on the dorsal tongue surface.
As the name implies, they are fungus-like or mushroom shaped structures.
They have thin non-keratinized epithelium and richly vascular connective tissue core.
They rise above the general level of the filiform papillae, and they usually have taste buds.
The taste buds are sparse in horse and cattle and abundant in carnivores and goats. They are found in the upper epithelial surface.
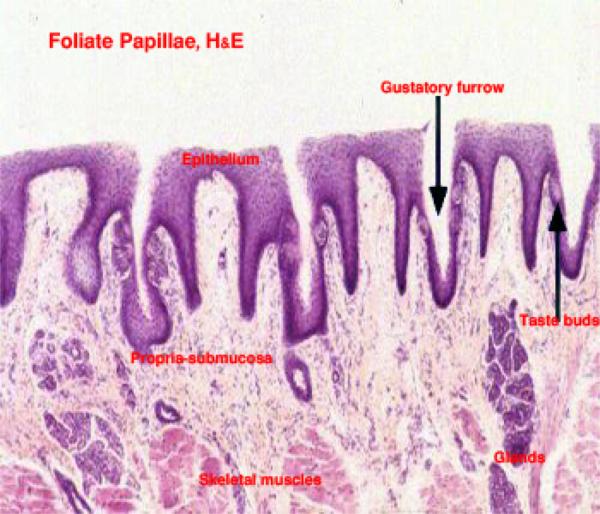
Foliate Papillae
The foliate papillae are parallel, leaflike mucosal projections located in the lateral borders of the tongue root.
The taste buds are located in the epithelium forming the sides of the papilla.
The foliate papilla is best seen in rabbits. ![]()
They are absent in ruminants and rudimentary without taste buds in cats.
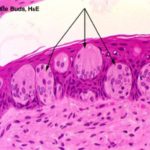
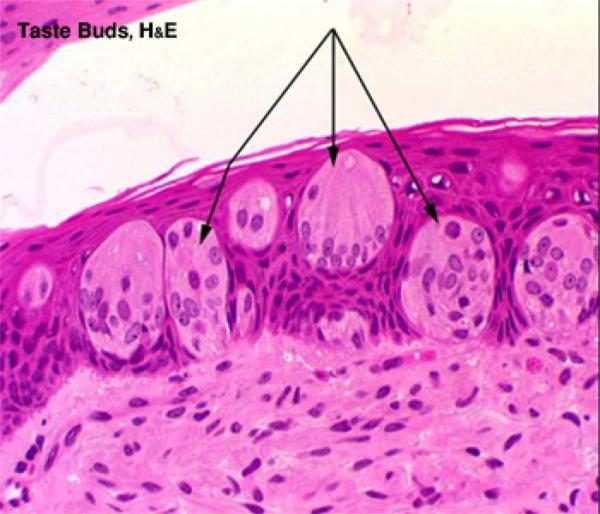
Taste Buds
The taste buds are specialized sensory organs embedded within the epithelium of the gustatory papillae.
They are also found scattered within the epithelial lining of the throat, soft palate and epiglottis.
With LM, they appear as oval-shaped clusters of pale-stained spindle-shaped cells that occupy the full thickness of the epithelium. ![]()
The cells forming the taste buds are of three types: gustatory, sustentacular or supporting and basal cells.
The gustatory, taste or sensory cells are located in the center of the taste buds.
Each cell bears a number of microvilli or taste hairs that project into a small opening within the luminal surface of the epithelium called taste pore.
With EM, their cytoplasm is rich in synaptic vesicles and they are associated with many afferent nerve fibers.
The supporting or sustentacular cells are spindle-shaped with much darker cytoplasm and are found among the taste cells.
The basal cells are population of small spherical cells located at the base of each taste bud and act as progenitors for other cell types.
Special structure of the tongue
The tongue of carnivore is characterized by the presence of a structure called lyssa.
The lyssa is a cord like structure extends longitudinally near the mid-ventral region of the tongue.
The lyssa is enclosed in a capsule of dense irregular collagenous fibers.
In dogs, the core of the lyssa is filled with adipose tissue, striated muscle fibers, blood vessels and nerves.
In cat, it contains mainly fat. The tongue of pig has a structure similar to lyssa.
The tongue of the horse has a mid-dorsal fibroelastic cord with hyaline cartilage, striated muscle fibers and fat.
Teeth
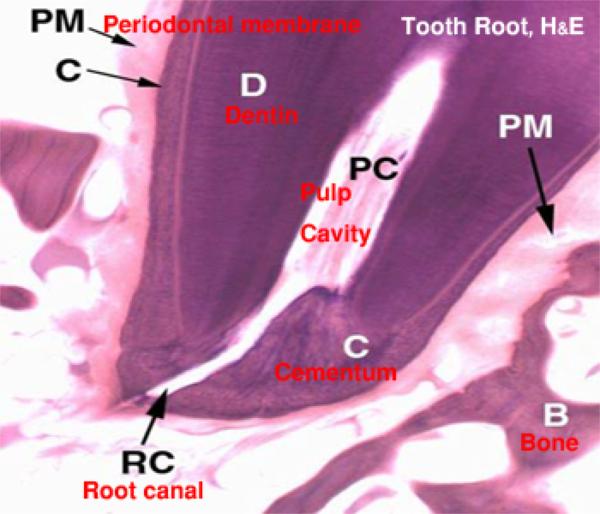
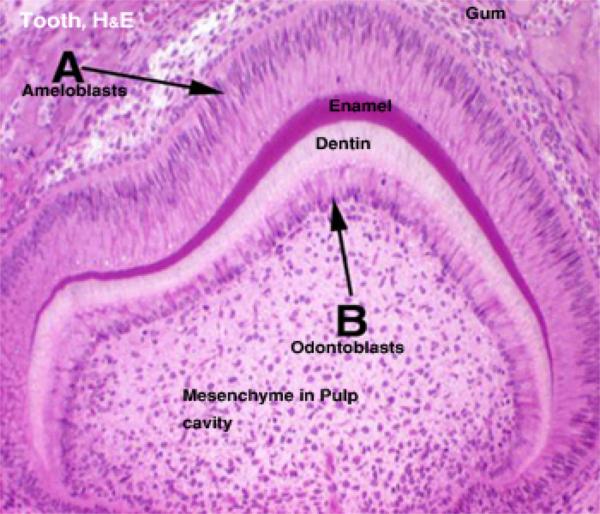
The tooth is grossly divided into two segments, the crown and the root: the crown is that portion which projects into the oral cavity and is protected by a layer of highly mineralized enamel.
The bulk of the tooth consists of dentine, a mineralized tissue which has a similar chemical composition to bone. ![]()
The dentine has a central pulp cavity containing the dental pulp which consists of specialized connective tissue containing many sensory nerve fibers.
The tooth root is embedded in a socket of jawbone, the bony alveolus.
The root of the bone is invested by a thin layer of cementum which is connected to the bone of the socket by a thin layer called the periodontal membrane.
The tooth is developing in a socket of the jawbone, the bony alveolus.
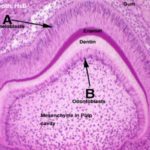
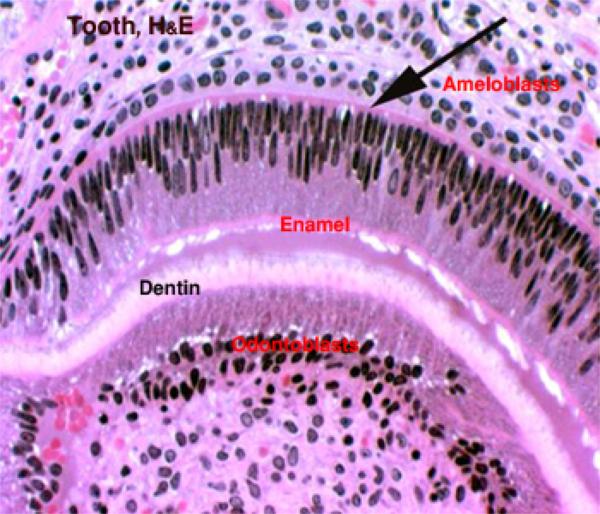
Primitive connective tissue, or mesenchyme has condensed in the future central region of the tooth, in what will become the pulp cavity. Inside this region has differentiated a population of specialized cells, the odontoblasts. ![]()
The odontoblasts make the bulk of the tooth’s hard material. They are visible as tall columnar cells, ranged side by side on the inside surface, closely applied to the wall.
Odontoblasts make dentin, by laying down a matrix material and then calcifying it.As they do so, they move backwards towards the center of the pulp cavity. This process continues after eruption, and as the animal ages the pulp cavity slowly diminishes in size.
On the outside surface of the tooth, a similar process is occurring; you will easily see a row of nicely columnar cells facing the odontoblasts. In this region enamel is being made by ameloblasts ![]() . As with the manufacture of dentin, ameloblasts also lay down an uncalcified matrix first, then harden it.Enamel is made only before eruption. Once the tooth erupts, the ameloblasts die, and if it’s damaged or chipped, it can’t be renewed.In some animals, especially in rodents, a portion of the enamel making apparatus remains viable below the gumline, and in this new enamel is made continuously.
. As with the manufacture of dentin, ameloblasts also lay down an uncalcified matrix first, then harden it.Enamel is made only before eruption. Once the tooth erupts, the ameloblasts die, and if it’s damaged or chipped, it can’t be renewed.In some animals, especially in rodents, a portion of the enamel making apparatus remains viable below the gumline, and in this new enamel is made continuously.
General Structure of Tubular Organs
The digestive tract has a distinct structural pattern that typifies tubular organs in general.
Although there are some minor variations from place to place, its structure usually includes four “tunics” or layers, a couple of which have subdivisions.
From the innermost (i.e., closest to the lumen) to the outermost, these are:
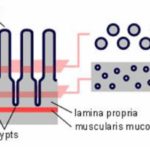
The tunica mucosa (which has subdivisions: epithelium, lamina propria and lamina muscularis mucosa);
The tunica submucosa (connective tissue layers with blood vessels, lymphatics and nerve plexuses),
The tunica muscularis (usually inner circular and outer longitudinal layers of smooth myscles with nerve plexus in between).
and an outermost tunica adventitia or serosa (connective tissue layer without (adventitia) or with (serosa) mesothelial covering.
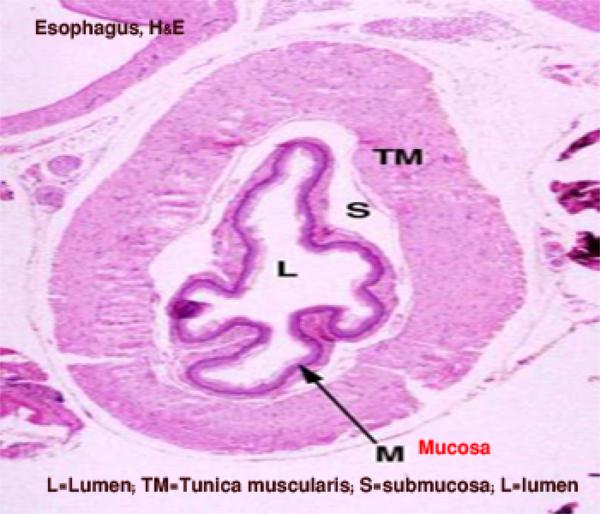
Esophagus
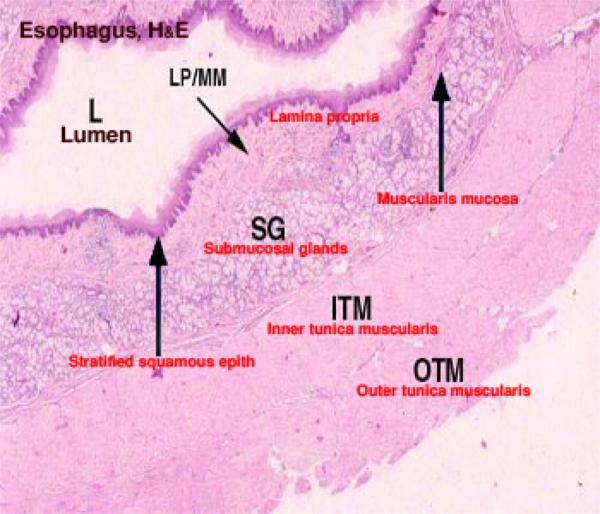
The esophageal wall is formed of mucosa, submucosa, tunica muscularis and serosa or adventitia.
The mucosa is formed of epithelium, lamina propria and lamina muscularis mucosa.
The epithelium is stratified squamous non-keratinised (dog, cat) slightly keratinized (pig) and highly keratinized (ruminants). ![]()
The lamina propria is a dense irregular connective tissue layer rich in immunocompetent cells, blood vessels and nerves.
The lamina muscularis is formed of longitudinally arranged smooth muscles.
In pig and dog, it is absent in the cranial end.
In cat, horse and ruminants, it is formed of isolated bundles at the cranial end and increase in number in the caudal part.
The submucosa is a loose connective tissue layer contains blood vessels, lymphatic, nerves and seromucoid esophageal glands. ![]()
In dog, the glands extend throughout the entire length.
In pig, they occur only in the cranial half.
In cat, horse and ruminants, they occur at the pharyngeo-esophageal Junction.
The tunica muscularis is formed of smooth and /or striated muscle fibers.
In dog and ruminants, it is entirely striated muscle.
In horse, it is striated in the cranial 2/3 and smooth in the caudal third.
In pig, it is striated in the cranial 1/3, mixed in the middle 1/3 and smooth in the caudal third.
In cat, it is striated in the cranial 4/5 and smooth in the caudal 1/5.
At the cranial end, the muscles are spirally arranged while caudally, an inner circular and an outer longitudinal layer are clear.
The cervical esophagus is covered by adventitia that is a loose connective tissue layer contains blood & lymph vessels and nerves.
The thoracic esophagus is covered by serosa that is a loose connective tissue layer with an outer mesothelial covering.
The abdominal esophagus (beyond the diaphragm) is about 2.5 cm in horse and short- wedge-shaped in carnivores is covered by serosa.
Glandular Stomach
The lumen of the glandular stomach is lined with simple columnar mucus-secreting epithelium.
There are no goblet cells in it (in which respect it differs from the intestines).
The mucosa is thrown up into longitudinal folds called rugae which stretch flat as the stomach distends.
If you were somehow able to stand inside the stomach and look at it en face, you’d see deep depressions in the “floor” representing gastric pits or foveolae, also lined with this simple columnar epithelium.
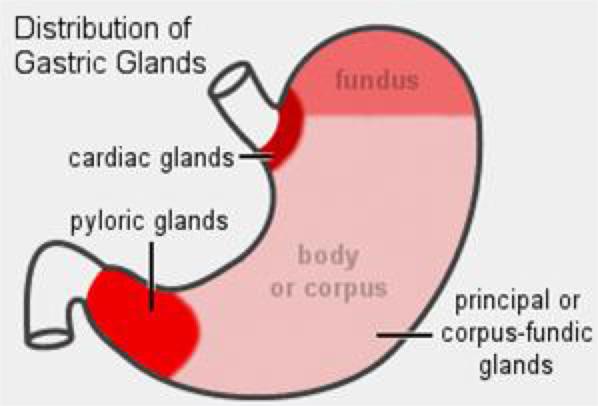
The lamina propria is a loose connective tissue layer rich in capillaries and lymphoid cells and is entirely occupied by glands, the gastric glands.
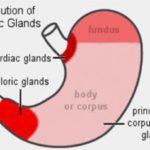
There are three major categories of these glands, associated with different parts of the stomach: cardiac, fundic and pyloric regions. ![]()
The glandular regions are wholly confined to the tunica mucosa.
Unlike some other portions of the tract, submucosal glands are not found in the stomach at all.
Cardiac Region
The first category is the cardiac glands.
These are found in the most proximal region of the stomach (i.e., that part closest to the input of the esophagus), called the “cardiac region,” because it’s located close to the heart.
The gastric pits are wide and deep.
At the bottom of the gastric pits are the openings into the secretory portions of the glands.
The glands themselves are short, branched tubular, slightly coiled with wide lumen extending deep into the mucosa, and are lined with simple cuboidal mucous secreting cells.
They are limited in their distribution to the region immediately adjacent the esophagus. Occasionally similar glands can be found in the esophagus itself. These are called “esophageal cardiac glands,” and probably represent individual variations in development.
Fundic Region
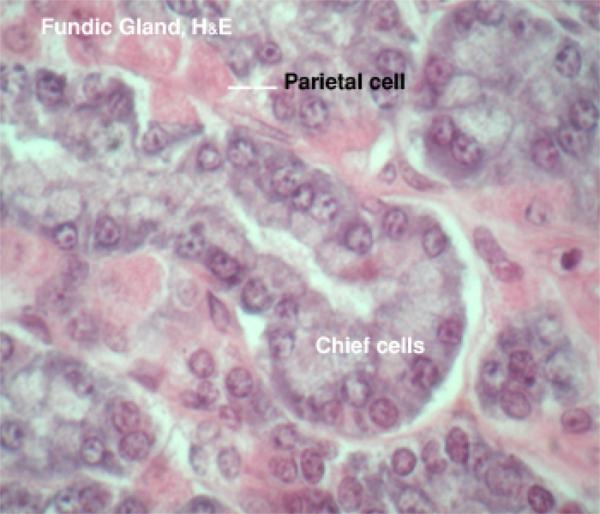
The second and by far the most numerous type of gastric gland is the fundic gland.
These are found underlying the bulk of the gastric mucosa.
These glands produce the bulk of secretions in the stomach.
They are deep, straight glands, with a mixed population of cells.
The fundic glands, like the other types, open into the base of comparatively shallow gastric pits.
Each gland consists of a neck, a long body and a dilated base or fundus.
The fundic glands are lined by four cell types; mucous neck cells, enzyme-producing cells, acid-producing cells and scattered endocrine cells.
Mucous Neck Cells
They are found in the neck of the gland and are smaller than the surface mucous cells.
They have basal nuclei and finely granular cytoplasm due to the presence of small mucus vacuoles that are distributed throughout the cytoplasm.
By LM, mucin granules do not appear unless stained by PAS.
Enzyme-Producing Cells
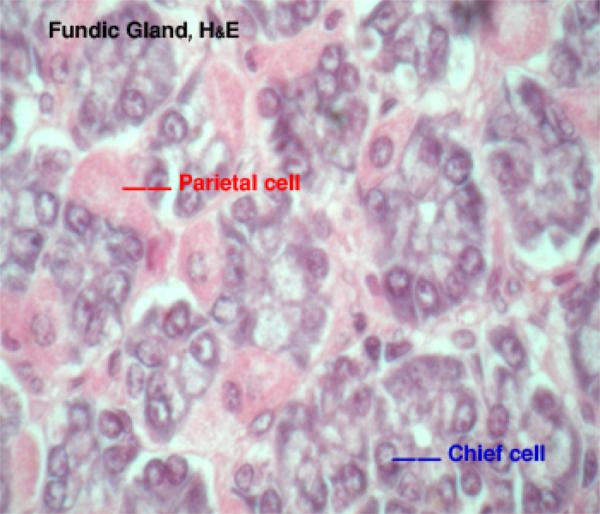
They are also called chief or peptic or zymogen cells. ![]()
They are the most numerous cell types within the fundic glands, hence the name chief cells.
The cells are cuboidal or pyramidal exhibiting all the criteria of protein-secreting cells: deep cytoplasmic basophilia, vesicular nucleus, and prominent nucleoli.
The cell apices appear acidophilic due to the presence of eosinophilic refractile cytoplasmic granules called zymogen granules. ![]()
With EM, the ultrastructure features are those of protein secreting cells; theses features include an extensive rER and membrane-bound secretory vesicles (zymogen granules) crowded in the apical cytoplasm, thus restricting the nucleus to the base of the cells.
The extensive rER accounts for the intense basophilia of peptic cells seen with LM.
The zymogen granules contain the inactive enzyme precursor pepsinogen, which is released into the lumen of the stomach where it is converted by the HCl into active enzyme pepsin.
Acid-Producing cells
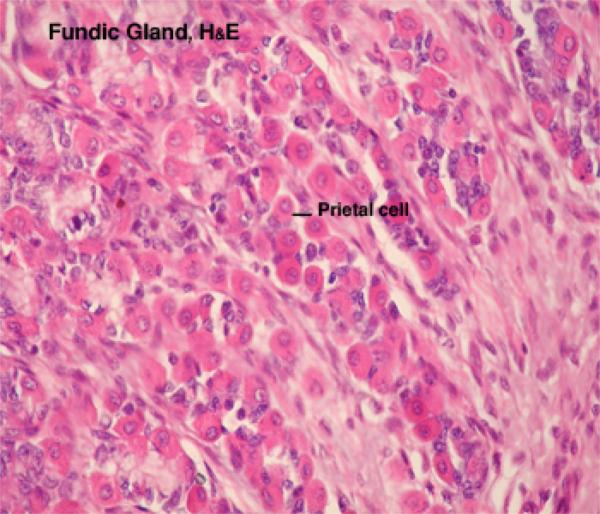
They are also called parietal or oxyntic cells.
They are round or pyramidal-shaped with spherical central nuclei. ![]()
They are very conspicuous, much larger than chief cells, and very strongly eosinophilic.
The base of the cell bulges outward and its narrow apex reaches the lumen of the gland.
With EM, the luminal plasma membrane forms deep, branching canaliculi which extend throughout the cytoplasm. Numerous short microvilli project into the lumina of the intracellular canaliculi. The cytoplasm is crowded with mitochondria as well as an extensive labyrinth of sER.
The parietal cells make hydrochloric acid, to keep the pH of stomach juice low (about 2.0 to 3.0 is typical). This pH is necessary to activate the gastric enzymes.
Parietal cells are also thought to secrete the substance called intrinsic factor which is essential for the absorption of vitamin B12 in the ileum.
Enteroendocrine
They are small spherical cells and are sited on the epithelial basement membrane.
In H&E sections, they have a spherical, central dark-staining nucleus and a rim of clear cytoplasm.
Some cells have an affinity for silver stains and are called argyrophil cells, others have an affinity for bichromate salts and are called chromaffin cells.
With EM, the cytoplasm contains membrane-bound neurosecretory granules.
The cells store and secrete serotonin, somatostatin, vasointestinal polypeptide-like (VIP), gastrin, bombesin-like peptides.
These hormones regulate the gastrointetinal functions.
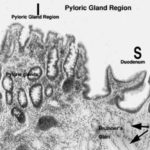
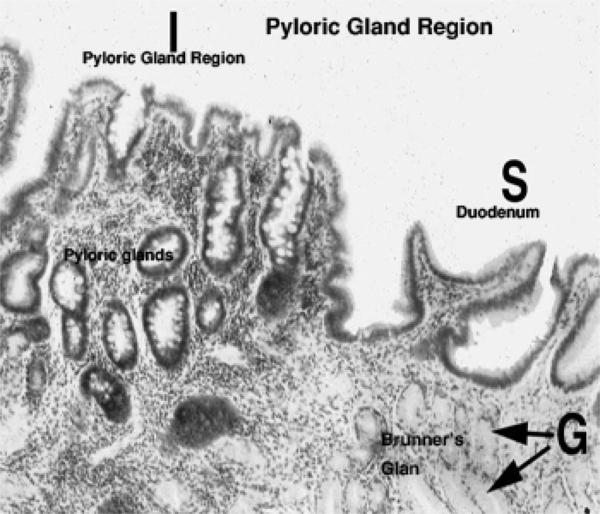
Pyloric Region
The third category of gastric glands is pyloric glands, found in the region of the pylorus, the junction with the first part of the intestine. ![]()
Pyloric glands produce a mucous secretion.
Structurally they closely resemble the glands of the cardiac region, though in the pyloric stomach the gastric pits tend to be deeper and the glands larger and more obvious.
The lamina muscularis of the stomach is made up of inner and outer circular and middle longitudinal layers of smooth muscle.
In the stomach of carnivores an additional layer, the subglandular layer, is located between the base of the gastric glands and the underlying lamina muscularis. It is made up of an inner stratum granulosum (fibroblast-rich) and an outer stratum compactum of dense collagen fibers.
The tunica muscularis of the stomach, unlike the bulk of the tract, is generally considered to have three layers.
The innermost layer is obliquely oriented with respect to the long axis of the organ.
There is a circularly oriented layer next outward of that, and the outer layer is longitudinal.
This arrangement of muscle causes the stomach to “wring” itself and to contract in length and diameter. The effect is to squish the softening food bolus into a liquid mass.
The outer surface of the stomach is covered by serosa consisting of loose connective tissue containing blood vessels, lymphatics and nerves and a single layer of mesothelial cells demarcates its outer limit.
Non-Glandular Region
The non-glandular region is lined by stratified squamous keratinized epithelium derived from that of the esophagus.
It is entirely absent in carnivores, small in pigs, wide in horse and reaches its greatest development in ruminants where it is subdivided into three distinct compartments; rumen, reticulum and omasum.
The structure of the other layers within the non-glandular are the same as seen for any tubular organ within the digestive tract.
Ruminants Stomach
The four divisions of the ruminant stomach are the rumen, the reticulum, the omasum, and the abomasum.
The first three compartments are referred to as forestomach. The fourth compartment is referred to as true stomach.
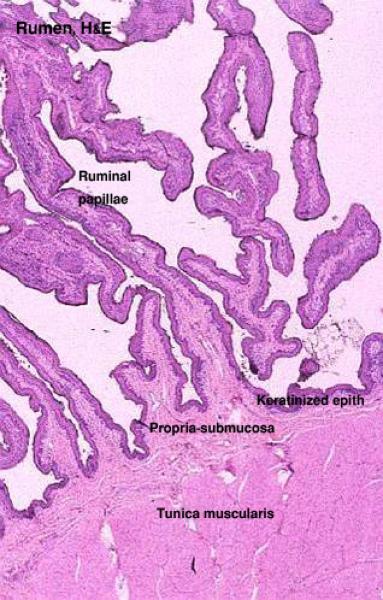
Rumen
The rumen is the site for microbial fermentation of fibrous ingesta. ![]()
Its mucosa is active in the absorption of short-chain volatile fatty acids produced during the fermentation process.
The ruminal mucosa has numerous finger-like or tongue-shaped projections called ruminal papillae.
The mucosa comprises lamina epithelialis and lamina propria.
The epithelium is stratified squamous keratinized epithelium.
The ruminal epithelium has protective, metabolic and absorptive functions.
There is no muscularis mucosae in the rumen.
The lamina propria and the submucosa combine to form a single underlying connective tissue support for the epithelium.
A dense network of fenestrated capillaries lies beneath the epithelium.
The submucosa is looser than the propria and contains many blood vessels, lymphatics and nerve plexus.
There are no glands present in either the propria or submucosa.
The tunica muscularis consists of inner circular and outer longitudinal layers of smooth muscles with myentric plexus in-between.
The serosa consists of loose connective tissue with an outer mesothelial covering.
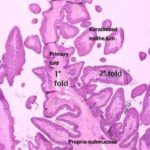
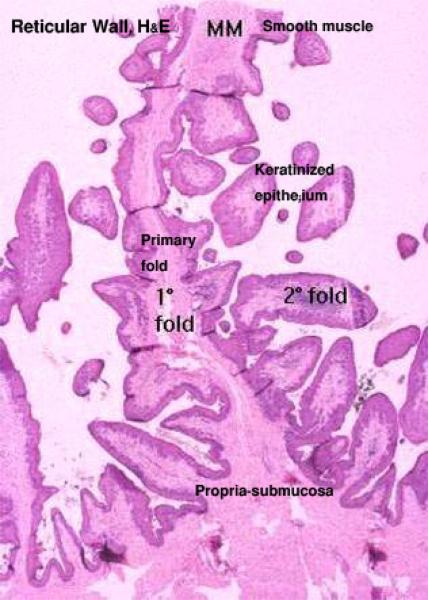
Reticulum
The reticulum is designed for mechanical breaking down of the ingesta into fine particles.
The reticular wall comprises mucosa, submucosa, tunica muscularis and serosa. ![]()
The reticular mucosa is organized into permanent anastomosing folds giving it a characteristic honeycomb appearance.
The sides of the folds have many conical papillae.
The lamina epithelialis is keratinized stratified squamous epithelium.
The lamina propria consists of a network of collagen and elastic fibers and there are no glandular elements.
A band of smooth muscles is located in the upper part of the large reticular folds derived from the lamina muscularis of the esophagus.
At places where the reticular folds intersect, the muscle bundles pass from one fold into another, forming a continuous network of smooth muscles throughout the reticular mucosa.
The contraction of the muscle bundles serves to contract the openings of the honeycomb, somewhat like a purse-string closes a purse.
The submucosa blends with the overlaying lamina propria without any demarcation, and is composed of loose connective tissue contains blood vessels, lymphatics and nerves.
The tunica muscularis consists of two oblique layers of smooth muscles with myentric plexus in-between.
The outer serosa is a loose connective tissue with an outer mesothelial covering.
Reticular sulcus
The reticular sulcus (esophageal groove or gastric groove) is well developed in milk-fed animals.
It begins at the cardia and ends at the reticulo-omasal orifice.
It is located on the medial wall of the reticulum and is bordered by two lips.
Contraction of the smooth muscle of the lips transforms the sulcus into channel that direct ingested milk and fluids from the esophagus directly to the abomasum by-passing the rumen and the reticulum.
The mucosa in the region of the reticular sulcus is covered by stratified squamous keratinized epithelium.
The lamina propria is made up of dense collagen and elastic fibers.
The lamina muscularis is a discontinuous layer of smooth muscle extended from the lamina muscularis mucosae of the esophagus, and is clearer in the lips.
The submucosa is a loose connective tissue layer.
The tunica muscularis consists of two layers: the inner layer is formed of transverse smooth muscle that extends into the lips. The outer layer contains both smooth and skeletal muscle fibers.
The serosa is a loose connective tissue layer with an outer mesothelial covering.
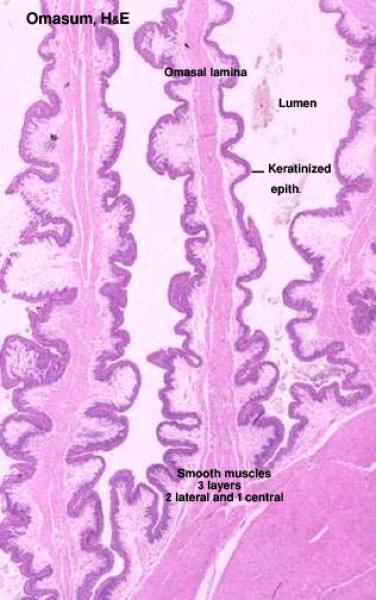
Omasum
The omasum is involved in mechanical breaking down of the ingesta into fine particles. ![]()
The mucosa is organized into longitudinal omasal laminae of five different lengths.
The lamina epithelialis is stratified squamous keratinized type.
The lamina propria is a dense irregular connective tissue layer rich in blood capillaries.
The lamina muscularis is made up of smooth muscle layers separating the lamina propria from the submucosa and forms two lateral bands inside each omasal lamina.
The submucosa is a loose connective tissue layer.
The tunica muscularis is formed of an inner circular and an outer longitudinal layers of smooth muscle fibers. The inner circular layer is extended into the large omasal folds as a central muscle sheet.
N.B. Each large omasal fold has, therefore, three muscle layers: two lateral layers derived from the muscularis mucosae, and one central layer originating from the inner circular layer of the tunica muscularis.
The outer serosa is a loose connective tissue layer with an outer mesothelial covering.
Abomasum
The abomasum is the true glandular stomach of ruminants.
Its histology is very similar to the fundic region of the stomach of monogastric animals.
The submucosa of this region is much more developed than in the previous divisions of the stomach.
The abomasum, unlike the rumen, reticulum, and omasum, does not absorb nutrients.
As with monogastric animals, it prepares food for enzymatic breakdown and absorption in the small intestine.
Small Intestine
The small intestine is the part of the digestive system responsible for the absorption of nutrients, but they could not function properly without the pancreas, which produces the digestive enzymes used for the process.
The small intestine is the main site for the absorption of amino acids, glucose, fat and some large molecules.
The intestines represent a nice application of a general rule in anatomy. If you want to improve the efficiency of a process which is associated with or limited by a transmembrane phenomenon, the way to do it is increase the surface area.
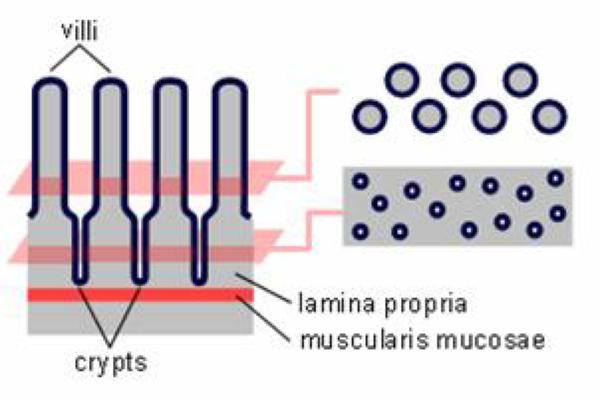
The small intestine has several modifications to increase its surface area:
First, the mucosa and submucosa are thrown up into a large number of folds or plicae arranged circularly around the lumen (plicae circularis).
Second, the surface of the plicae is further arranged into villi ![]() that protrude into the intestinal lumen.
that protrude into the intestinal lumen.
Third, the absorptive surface is further increased by the presence of microvilli on the free surface of the absorptive epithelial cells.
Fourth, the intestinal mucosa is invaginatd between the bases of the villi into crypts called crypts of Liberkühn. ![]()
The small intestine is subdivided histologically into three distinct regions: duodenum, jejunum and ileum.
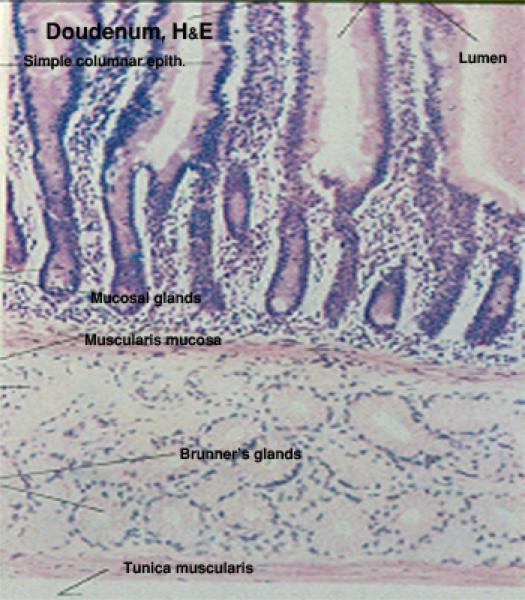
Duodenum
The duodenum is the first portion of the small intestine.
The epithelium of the duodenal mucosa is a regular simple columnar form, and the microvilli of these enterocytes form a continuous brush border.
The brush border is visible under high magnification as a refractile band along the apical surface of the cells.
The duodenal mucosa is arranged in the form of short, uniformly sized villi, all quite regular in size and shape.
Between the villi are the intestinal crypts which extend downward to the deepest levels of the tunica mucosa.
The intestinal crypts are simple straight tubular glands lined by enterocytes, goblet cells, stem cell, Paneth cells and enteroendocrine cells.
Most of the epithelial cells are the absorptive enterocytes.
The goblet cells are glass shaped unicellular glands that secrete a mucoid material which coats and protects the enterocytes.
The enteroendocrine cells are less numerous.
They are similar to those in the stomach, but producing different hormones.
They are located in the deep regions of the crypts.
They have granules in their cytoplasm, in which the granules are polarized away from the lumen of the crypt and towards the lamina propria.
These cells don’t secrete into the lumen, they discharge their secretions into the blood flowing through capillaries in the lamina propria.
The Paneth cells are found in clusters at the bases of the crypts in ruminants, horse and man.
The cells are pyramidal with basal spherical nuclei.
The luminal cytoplasm contains prominent large eosinophilic granules.
With EM, the granules are spherical and electron-dense, the remaining cytoplasm is rich in rER (protein-secreting cells).
They are believed to produce peptidase and lysozyme which are bactericidal materials and release them into the lumen of the gut.
Note that the Paneth cells aren’t part of the enteroendocrine system because they don’t release hormones into the blood, they release it into the gut lumen.
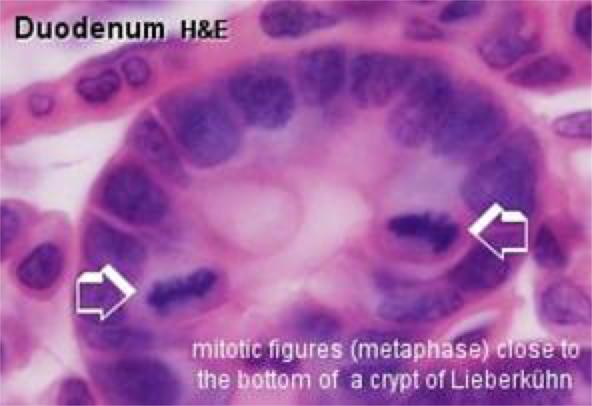
The stem cells are found in the lower third of the crypts. ![]()
They divide to replace the enterocytes and mucous cells of the villi since these cells have a rapid turnover (about 5 days).
Paneth cells and endocrine are also replaced by the stem cells.
Unlike the stomach, the crypts (which are called “glands” in some texts) do not secrete digestive enzymes or hydrochloric acid.
The enterocytes rest on a thin basal lamina and immediately below is a network of fine collagenous and elastic fibrils, heavily infiltrated with cells, which is the lamina propria.
The lamina propria is a loose and very cellular irregular connective tissue. Most of the cells within the meshes of the collagen fibrils are plasma cells, although many other cell types (including almost all of the true cells of the blood) can be found.
Within the lamina propria, and up into the core of the villi there are mall strands of smooth muscle, which come off the muscularis mucosae.
The core of each villus has numerous small capillaries in its lamina propria, and a single large lymphatic vessel, called the central lacteal. This is a blind-ended structure which represents the beginning of the intestine’s lymphatic drainage. ![]()
The lamina propria is demarcated from the more peripheral layers by the muscularis mucosae, a thin band of smooth muscle which runs all the way around the mucosal layer and which sends strands up into the villi.
It’s believed these strands cause the villi to contract, expelling the contents of the crypts and the intervillous spaces.
The tunica submucosa is the region of connective tissue immediately outside the muscularis mucosae. It has fair numbers of blood vessels and lymphatics as well as collections of neuron cell bodies. These are elements of the submucosal plexus This plexus, together with another one located in the tunica muscularis helps to coordinate the movements of the intestine and facilitate the passage of food through its lumen.
The submucosa of the duodenum also has true glandular elements, called Brunner’s glands. ![]()
These submucosal glands are specifically a feature of the duodenum, and are typically found in the first portion of it. They aren’t present in the jejunum or the ileum.
The submucosal glands produce an alkaline secretion which helps to neutralize the very acidic (pH 2.0-3.0) material entering the duodenum from the pyloric region of the stomach.
The tunica muscularis is quite thick, and in the duodenum (as well as in the rest of the intestines) has the typical two layers, inner circular and outer longitudinal.
Between these two layers is the second nervous plexus, of the intestine, the myenteric plexus (formerly called the plexus of Auerbach).
The plexus appears as discrete areas of neuron cell bodies, located at fairly regular intervals between the inner and outer muscle coats.
The outer serosa is a loose connective tissue layer with an outer mesothelial covering.
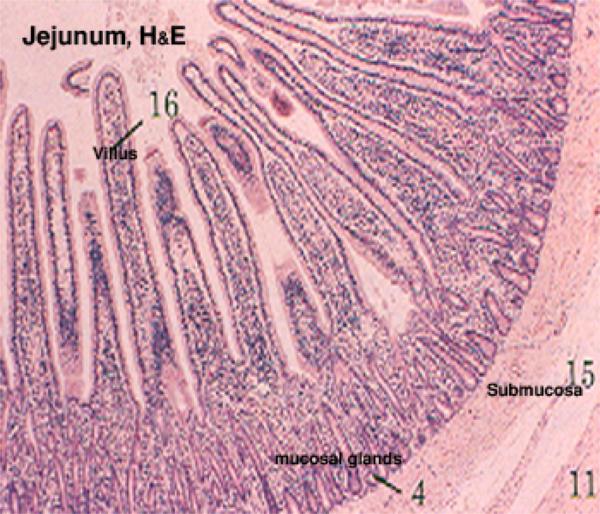
Jejunum
The next section of the small intestine is the jejunum.
Jejunal villi are longer and more irregular than the short, blunt villi characteristic of the duodenum.
It’s in this section that the bulk of absorption of nutrients takes place. ![]()
The muscularis mucosae here is sparse or even absent and there are gross folds which involve not only the tunica mucosa., but also the underlying submucosa.
These folds, the plicae circulares, are permanent structures, and another means for increasing surface area.
Other features are the same as those in the duodenum.
Notice that there are no glands in the submucosa in this region.
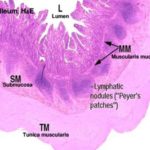
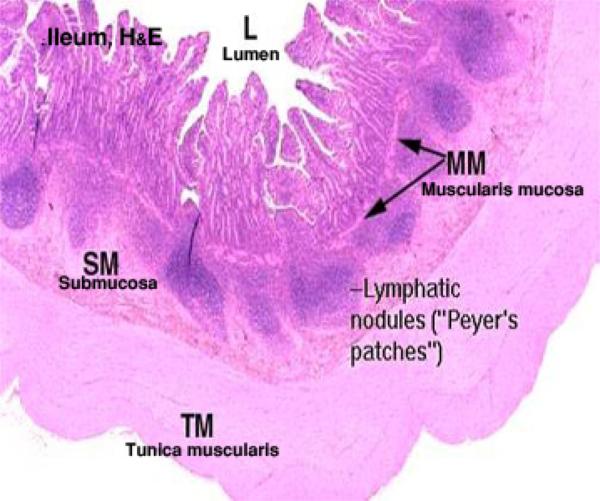
Ileum
The ileum is the last portion of the small intestine.
Most of the features you will see in this portion of the intestine are similar to those in the previous sections, but the villi are even more leaflike. ![]()
The most prominent landmark in the ileum is the presence of large aggregations of lymphatic tissue in the submucosa.
These are the aggregated lymphatic nodules (once called Peyer’s patches).
These are often so large that they will infiltrate past the muscularis mucosae and up into the mucosa proper, sometimes obliterating the enterocytes.
The submucosal lymphatic nodules frequently contain germinal centers, and they are an important part of the lymphatic system.
They represent the site of maturation and development of the B-lymphocytes important in humoral immunity.
While not transient structures, they do vary in number depending on the age of the animal, and germinal centers are more frequently seen in them in younger animals.
Large Intestine
The large intestine lies between the ileum and the anus.
It comprises the cecum, colon, rectum and anus.
The principal function of the large intestine are: reabsorption of water and soluble salts, bacterial degradation of cellulose in ruminants, absorption of fat soluble vitamins (A, D, E, and K) produced through bacterial activity, and elimination of wastes.
Between the ileum and the colon proper is a diverticulum, called the cecum. This is described in most texts as part of the colon, but it actually has openings into both the colon and the ileum in most species.
The layers of the cecum are similar to those of the small intestine, but note that villi are absent; there are only crypts.
The number of goblet cells has increased, there is a great deal of fat in the connective tissue of the submucosa, and the tunica muscularis is much scantier.
There are some small lymphatic nodules present.
The histology of the cecum is very similar to that of the colon.
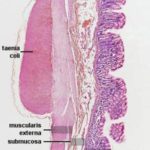
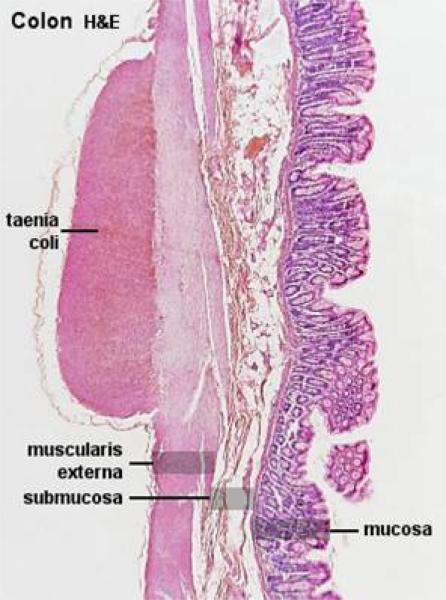
Colon
The mucosal layers are similar in arrangement and histology to the cecum.
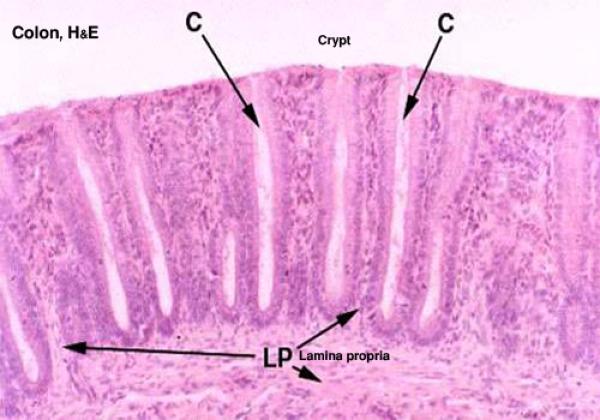
The mucosa of the colon is smooth and devoid of villi. ![]()
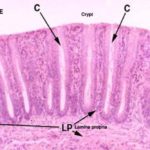
The epithelium is simple columnar with an increase number of goblet cells.
The lamina propria is formed of loose lymphoreticular tissue entirely occupied by intestinal crypts. Solitary lymph nodules are found between the crypts.
The intestinal glands or crypts of Leiberkühn are simple straight tubular downgrowths extending from the surface to the muscularis mucosae. ![]()
They are lined by columnar cells, abundant goblet cells, stem cells, enteroendocrine cells with NO Paneth cells.
The submucosa is devoid of glands except in some parts of the colon.
The tunica muscularis is formed of the usual inner circular and outer longitudinal layers with myentric plexux in-between.
In the colon, the outer layer is not continuous, being concentrated into three bands, the taeniae coli.
The outer serosa has the usual structure.
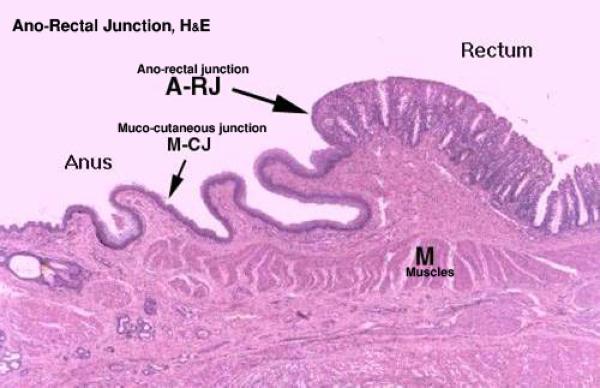
Ano-Rectal Junction
At the anorectal junction, the epithelium changes abruptly from simple columnar to stratified squamous non-keratinized. ![]()
The lamina propria-submucosa is gland free (ruminants and horse).
In dog, cat, it contains modified tubulo-alveolar apocrine sweat glands as well as sebaceous like glands (in cats only) that open in anal sinuses by ducts.
The secretion of the sebaceous glands help to lubricate the duct of the anal glands thus guards against its obstruction.
The absence of sebaceous glands in dogs explains the frequent obstruction of the ducts of the anal glands, which is a common clinical finding in this animal.
The ducts and sac are lined by stratified squamous epithelium and opens into the anus at the level of anocurtaneous junction.
The lamina muscularis and the outer longitudinal layer are absent. The inner circular layer forms the internal sphincter. The external anal sphincter composed of circular skeletal muscle.
Accessory Glands
The accessory glands include the liver, the pancreas, and the salivary glands.
Liver
The liver is the largest gland in the body.Its major important functions are detoxification of metabolic waste products, destruction of spent red cells and recycling of their constituents, synthesis and secretion of bile, synthesis of plasma proteins including the clotting factors, synthesis of plasma lipoproteins and various other metabolic functions e.g., glycogen synthesis and gluconeogenesis.The liver is covered by a thin connective tissue capsule covered externally by mesothelium derived from the visceral peritoneum.The outer capsule sends trabeculae into the hepatic parenchyma dividing it into lobules.
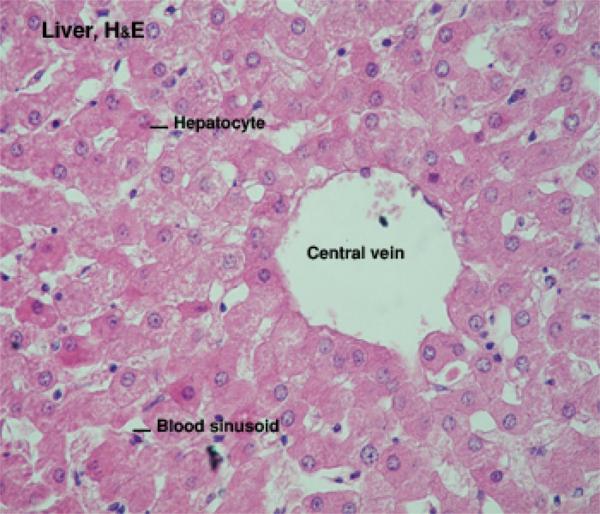
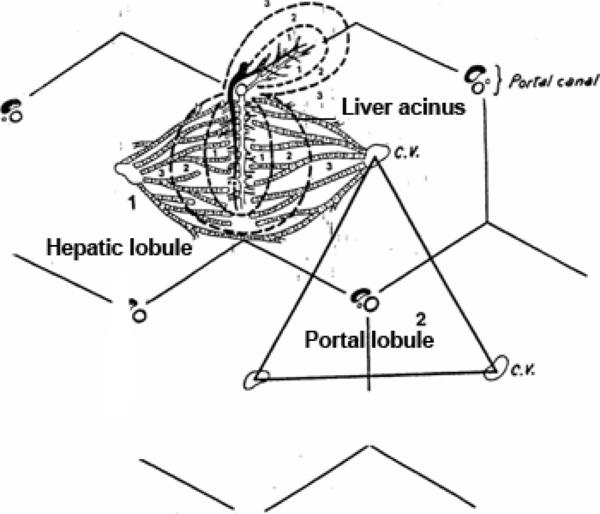
Hepatic Lobule
The classic hepatic lobule is considered to be the functional unit of the organ. ![]()
Each is a roughly hexagonally shaped, three dimensional unit, demarcated by connective tissue and constructed of the parenchymal cells of the liver, hepatocytes, in large numbers.
The pig’s and camel’s livers have more connective tissue than is true of most other animals; this abundance of connective tissue results in a very clear demarcation of hepatic lobes in these animals.
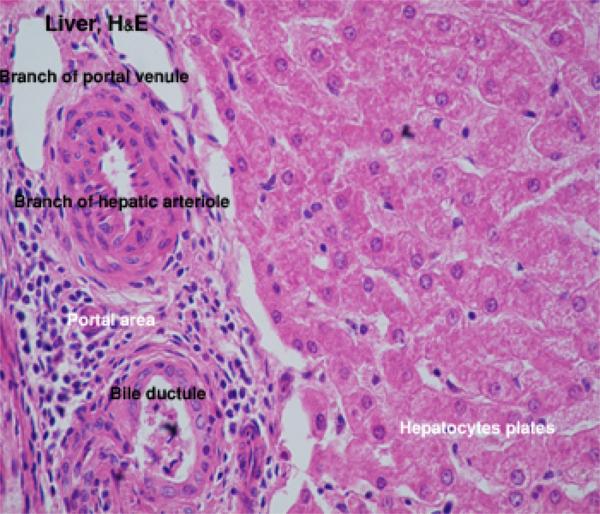
At each corner of the liver lobule, there are portal area or portal canal or hepatic triad.
The portal areas include at least one each of the following elements: 1) a branch of the hepatic artery; 2) a branch of the portal vein; and 3) a bile ductule. ![]()
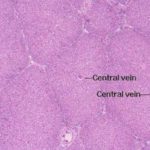
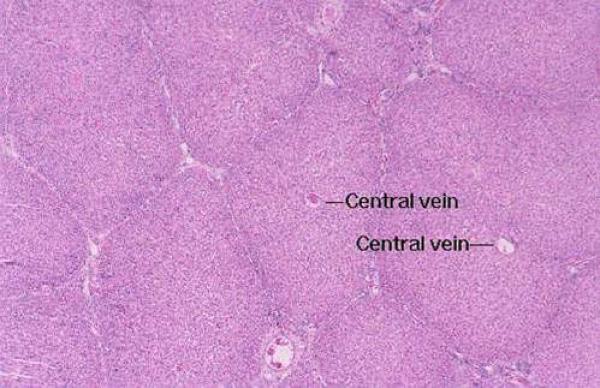
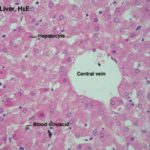
Within the limits that define each lobule, the hepatocytes are arrayed in long rows, the hepatic laminae and each lobule is organized around a so called central vein ![]() , really just an open space.
, really just an open space.
Each lobule is supplied by blood from two sources: a branch of the hepatic artery and a branch of the portal vein.
Blood coming in from the periphery flows through the sinusoids that separate the plates of liver cells, and eventually finds its way to the central vein.
The flow of blood is from the portal canal’s vascular elements into the sinusoids; through the sinusoids and into the central vein, which is the beginning of the organ’s venous outflow.
The drainage from each lobule eventually travels through larger and larger vessels, and is merged with the outflow from others.
Finally all of the blood leaves the liver through the hepatic vein, which enters the right side circulation.
Most of the blood which enters the organ comes in via the portal vein, which drains capillary beds in the intestine; a much smaller amount comes in via the hepatic artery. Thus, both oxygenated and deoxygenated blood are mixed in the sinusoids.
The hepatocytes are large polyhedral cells fairly uniform in size, which have variable cytoplasmic appearance depending on the nutritional status of the body.
In well-nourished individual, their cytoplasm store significant quantities of glycogen and lipids hence appear vacuolated. The remaining cytoplasm is strongly eosinophilic due to a high content of organelles.
The nuclei are large with peripherally dispersed chromatin and prominent nucleoli. As many as 25% of hepatocytes are binucleated. More than half of the hepatocytes contains twice the normal (diploid) complement of chromasomes within a single nucleus (i.e., tetraploid). Some contains four or even eight time (i.e., polyploid).
With EM, the cytoplasm is crowded with organelles, particularly rough and smooth ER, mitochondria, and lysosomes. Lipid droplets and glycogen rosettes are present.
Plates of hepatocytes are radially arranged around the central vein.
These plates are usually one cell thick.
The hepatocytes make bile, sequester and release carbohydrates, and produce the amazing varieties of protein products.
The hepatocytes take up and store excess glucose as glycogen.
Glycogen can, when needed, be breaking down back into glucose by the hepatocytes, and put into the blood.
Hepatocytes are temporary storage depots for lipids.
The sinusoids are irregular vascular channels between the plates, with numerous blood cells in them.
A narrow space known as the space of Disse is located between the sinusoidal lining cells and the hepatocyte surface.
The sinusoidal lining cells are discontinuous and thus, the space of Dissue is continuous with the lumen of the sinusoids.
The hepatocyte surfaces facing the blood sinusoids carry numerous microvilli that extend into the space of Disse, thus increasing the surface area available for exchange of metabolites between liver and blood.
The Kupffer cells are phagocytic cells, derived (as are other examples of macrophages) from the monocyte cell line.
These liver-resident macrophages engulf particulate matter that passes through the sinusoids (especially bacteria and senescent blood cells).
They are a little difficult to make out, but you should be able to spot them by the presence of brownish pigment particles inside. This pigment is the product of cell breakdown and can be either lipofuscin or hemosiderin.
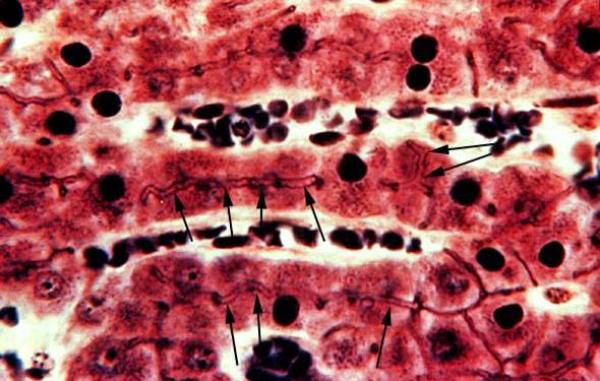
The bile canaliculi are minute channels forming an anastomosing network between hepatocytes.
The wall of the canaliculi is formed of the plasma membrane of adjacent hepatocytes. ![]()
Bile is synthesized by all hepatocytes and is secreted into the bile canaliculi that drain towards bile collecting vessels located in the portal areas.
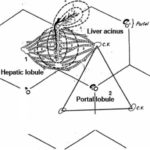
Portal Lobule
The portal lobule is the functional unit of the liver based on the exocrine activity.
It is triangular in shape and is made up of a portal tract and parts of the three adjacent hepatic lobules.
The boundaries of the portal lobule are defined by lines joining three adjacent central veins. ![]()
Bile secreted from the hepatocytes located inside the portal lobule is drained into bile ductule located in the central portal area.
Liver Acinus
The liver acinus is the functional unit of the liver based on the blood supply of the hepatic cells.
It is a diamond or oval-shaped structure formed by the hepatocytes of two adjacent hepatic lobules.
The margins of the diamond shape are drawn between two central veins. ![]()
The hepatocytes located inside the liver acinus are supplied by the same interlobular vessels. The liver acinus is subdivided into three zones: zone I, zone II and zone III. Zone I lies near the interlobular region and receives the best blood supply. Zone II is the intermediate zone and receives blood of moderate quality. Zone three lies close to the central vein and receives poor blood supply, therefore it is the first zone to die.
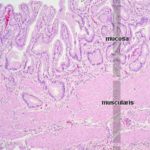
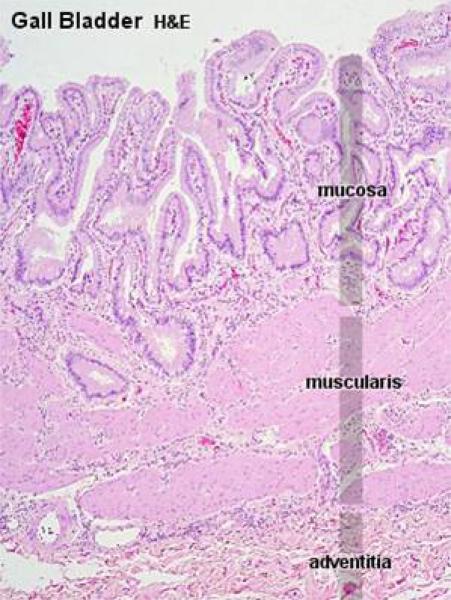
The Gall Bladder
The gall bladder is absent in rats, camels and horses as well as pigeon.
Its function is to concentrate and store bile produced in the live. ![]()
The wall of the gall bladder is formed of mucosa, submucosa, tunica muscularis and serosa.
The mucosa is covered by a very regular simple columnar epithelium, with no goblet cells or glands.
There is a scanty lamina propria.
There is no muscularis mucosa.
The lamina propria-submucosa is formed of loose connective tissue containing solitary lymph nodules.
The deep folds in the bottom of the mucosa, which are often cut in cross section, sometimes look like “glands,” but they have no secretory activity and they are really parts of the lumen.
In ruminants, true glands of seromucoid type are present.
The tunica muscularis is thin and is formed of circular layers of smooth muscle fibers.
The serosal covering (i.e., visceral peritoneum) covers the portion of the organ that’s not nestled up to the liver.
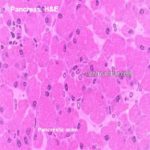
Pancreas
The pancreas synthesizes the bulk of the digestive enzymes needed in the intestines.
Impairment of this function will lead to maldigestion, malabsorption, and eventual death.
Inflammation and especially cancer of the pancreas are very serious and often fatal for this reason.
The pancreas is a highly lobulated compound tubuloacinar gland with both exocrine and endocrine portions; digestive enzymes are conducted via a duct system to the duodenum, and the endocrine products enter the blood directly.
The pancreas is invested by a loose connective tissue capsule which sends delicate septa demarcating the pancreas into lobules.
Each lobule is formed of exocrine serous secreting units, ducts and endoctine islets of Langerhans.
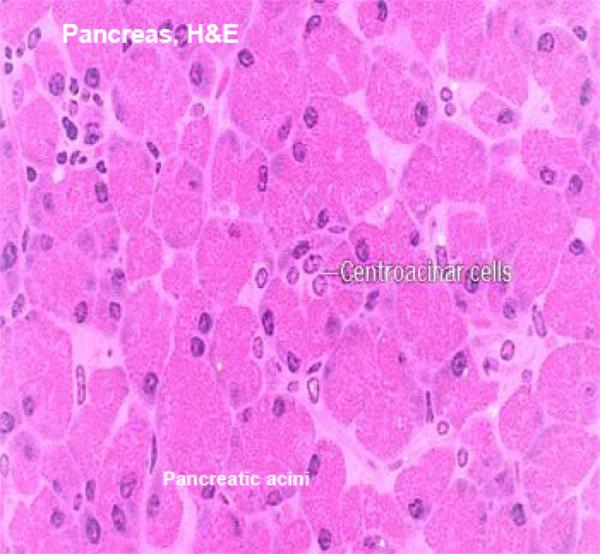
The bulk of the tissue in the pancreas is exocrine in nature. This is the pancreatic acinar tissue and its associated ducts. ![]()
Exocrine Pancreas
The cells of the acinar tissue are arranged into blind-ended acini (that’s a plural–the singular is “acinus”) as in other glands.
The pancreatic acini are formed of pyramidal-shaped cells with their apices projecting towards the lumen of a minute duct.
The lumen of an acinus is small.
The acinar cells are typical proteins secreting cells.
The nuclei are spherical basally located and surrounded by basophilic cytoplasm. The apical region contains distinct granular material, the zymogen granules.
The zymogen granules are eosinophilic and are precursor forms of digestive enzymes.
The deep basophilia of this region is due to the presence of large amounts of rough endoplasmic reticulum, which binds the hematoxylin of the H&E stain heavily.
The pancreatic duct begins from the lumen of each acinus.
The intercalated ducts are short ducts. Each duct drains only one acinus. The duct lining cells are often seen in the lumen or the center of secretory acini, hence they are called centro-acinar cells, and are very characteristic to pancreatic ducts.
The intercalated ducts of the adjacent acini merge with each other forming intralobular duct, so named because it is located inside the lobule. The pancreatic intralobular ducts are lined by simple cuboidal epithelium, and in contrast to intralobular ducts of the salivary glands they are not striated.
The intralobular ducts fuse with each other and now they become larger and extended up to the level of the interlobular septa. These are the interlobular ducts ![]() which are lined with simple columnar epithelium instead of the cuboidal one.
which are lined with simple columnar epithelium instead of the cuboidal one.
The secretions from the interlobular ducts are collected to a larger duct, the main pancreatic duct which drains into the duodenum.
The main duct is lined by simple columnar epithelium with some goblet cells.
The ducts in the pancreas are not so obvious as those in salivary glands, but the layout of the glands is similar.
Endocrine Pancreas
The endocrine portion is the pancreatic islets of Langerhans are regions of lighter staining tissue in the mass of the exocrine part. The islets are demarcated by a very fine investment of delicate CT fibrils. They’re well vascularized (as you would expect in any endocrine organ) The cells produce the hormones insulin and glucagon. The endocrine pancreas will be taken with more consideration on dealing with the endocrine organs.
Salivary glands
The salivary glands are glands that produce the saliva.
They are divided into two major groups; minor and major salivary glands.
The minor salivary glands are small clusters of seromucoid secretory units located in the submucosa of the oral cavity and they are named according to their locations e.g., labial, buccal, lingual and palatine glands.
The major salivary glands are located outside the oral cavity and their secretory products reach the oral cavity via excretory ducts. They include parotid, mandibular, sublingual, zygomatic (in carnivores) and molar (cat) glands.
General Structure of the Major Salivary Glands
The major salivary glands are compound tubuloacinar glands.
Each gland is surrounded by a capsule of dense irregular connective tissue and is divided into numerous lobules each containing many secretory units.
Connective tissue septa radiate between the lobules from the outer capsule and convey vessels, nerves and large excretory ducts.
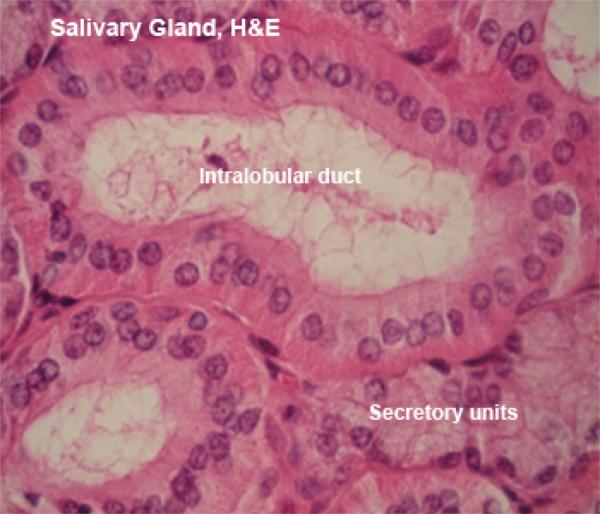
Th terminal secretory units merge to form the intercalated ducts which are lined by low cuboidal epithelium.
The intercalated ducts drain into larger ducts called striated ducts, so named because of their striated appearance in LM. Both the intercalated and striated ducts are located inside the lobule, hence they are collectively called intralobular ducts.
The striated ducts join the interlobular ducts which are located in the CT septa between the lobules. They are lined with simple columnar epithelium in the smaller ducts and with stratified columnar in the larger ones.
The main duct is lined by stratified columnar epithelium which changes into stratified squamous type as it opens into the oral cavity.
The salivary secretory units consist of branched tubuloacinar structures and are composed of either serous, mucous secretory cells or a mixture of both types.
Parotid Salivary Gland
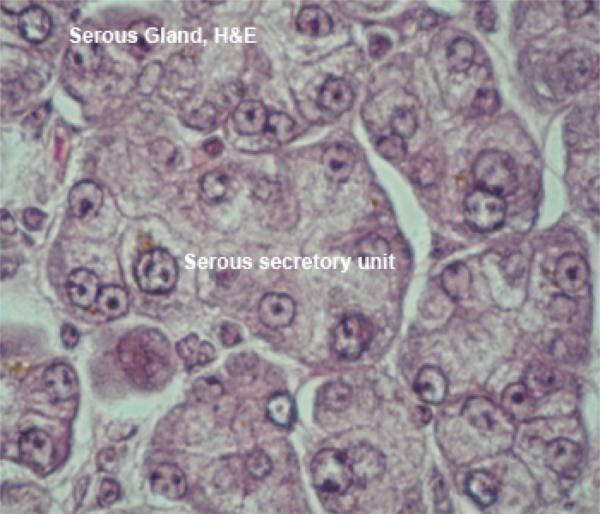
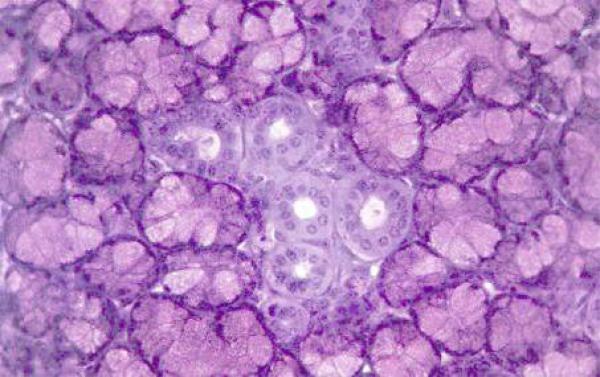
The parotid glands are large paired predominantly serous glands. ![]()
Isolated mucous secretory units may occur in dog and cat.
Mandibular Gland
It consists of serous and mucous cells but the serous cells predominate.
Mucous secretory units border the lumen and serous demilunes occur at the periphery.
Sublingual Gland
It is of mixed seromucoid type but the mucous cells predominate. ![]()
The striated ducts are not well-developed and the duct system is formed mainly of interlobular ducts.
Zygomatic Glands (Carnivores)
The gland is located beneath the zygomatic arch of the temporal bone.
It is of mixed seromucoid type but mucous units predominate.
The intercalated and striated ducts are almost non-existent. Isolated patches of tall striated cells are found within the low cuboidal epithelium of the interlobular ducts.
Molar Salivary Glands
The gland is located beneath the mucosa of the lower lips near the commissure.
It is of mixed seromucoid type but the mucous units predominate.
The intercalated and striated ducts are absent.