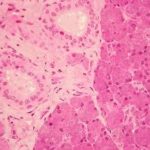
-
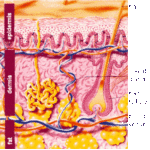
- Figure 1
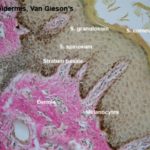
-
- Figure 2
-
- Figure 3
-
- Figure 4
-
- Figure 5
-
- Figure 6
-
- Figure 7
-

- Figure 8

-
- Figure 9
-
- Figure 10
-
- Figure 11
-
- Figure 12
-
- Figure 13
-
- Figure 14
-

- Figure 15
-
- Figure 16
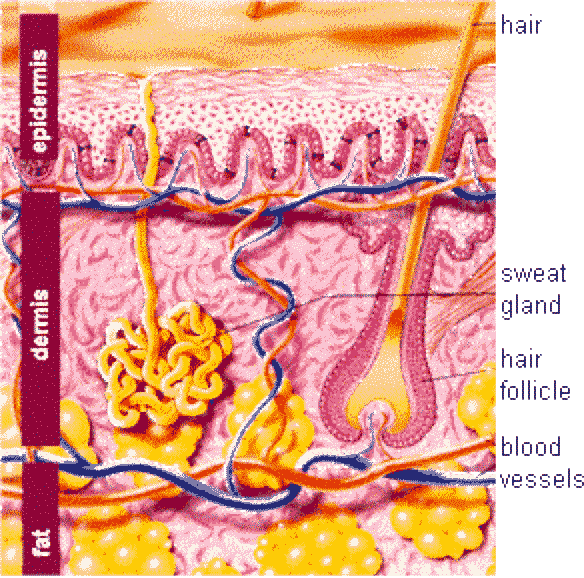
Integument (Skin)
The integument of mammals consists of the skin, and also the various accessory structures such as hair, glands, nails, hooves, claws.
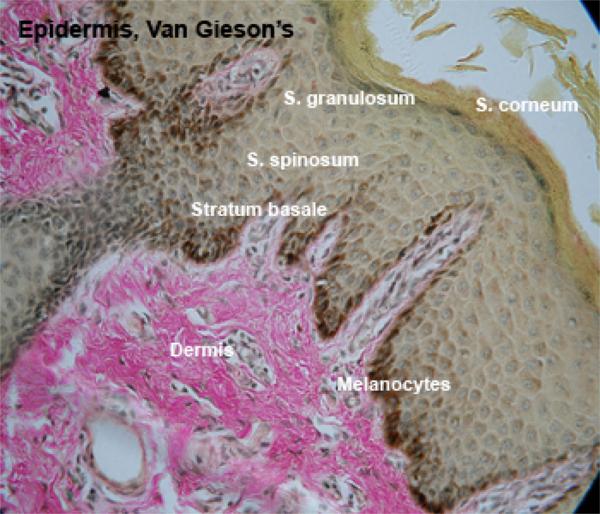
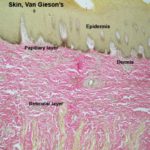
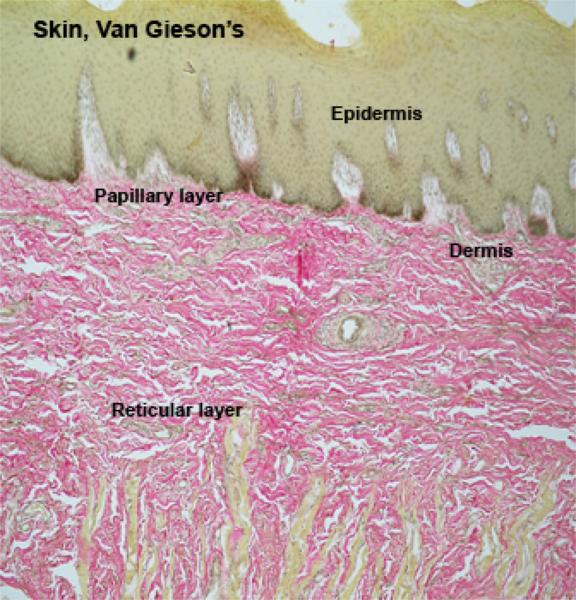
![]() The skin is formed of epidermis, dermis, and hypodermis.
The skin is formed of epidermis, dermis, and hypodermis.
The epidermis is a keratinized stratified squamous epithelium comprising the following layers: stratum basale, stratum spinosum, stratum granulosum, stratum lucidum, and strutum corneum.
The deepest layer of the epidermis is the stratum basale.
This layer is only one cell layer of cuboidal to columnar cells.
It lies rights up against the dermo-epidermal boundary.
It’s here that almost all of the proliferative activity occurs.
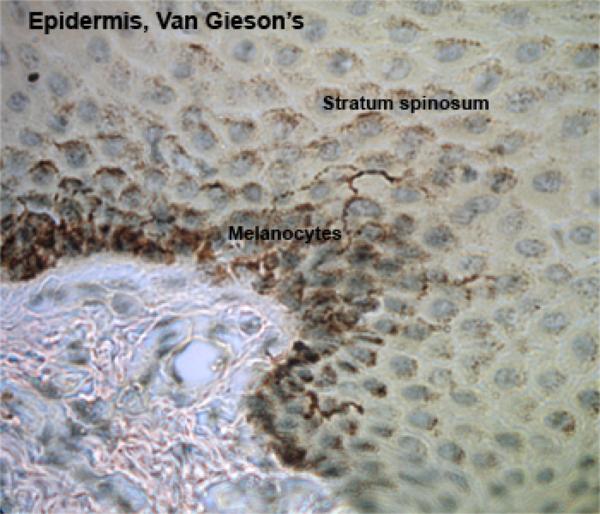
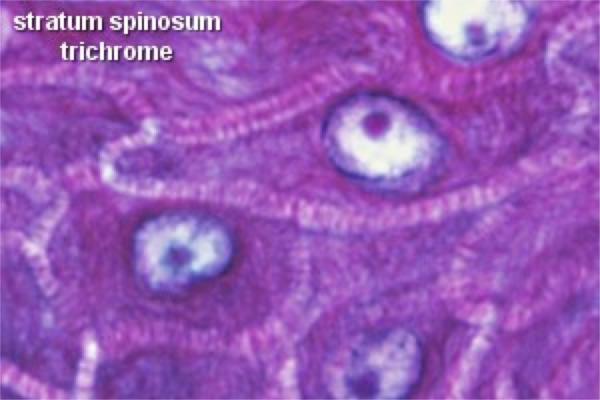
The stratum spinosum (prickle cell layer) consists of several layers of polyhedral cells fixed to each other by numerous desmosomes.
In this layer the cells are still alive, they have prominent nucleoli and cytoplasmic basophilia that indicate active protein synthesis.
The cells secrete fibrillar protein that aggregates to form intracellular fibrils known as tonofibrils, which converge upon the desmosomes.
The stratum spinosum may be quite extensive in thick skin, but in thin skin it may be only one or two cells breadth.
The stratum basale and the deep layer of the stratum spinosum show numerous mitotic figures, hence they are called stratum germinativum.
The stratum granulosum consists of few layers of flattened cells that accumulate dense basophilic keratohyaline granules in their cytoplasm.
The process of keratinization involves the combination of tonofibrils and keratohyaline granules to form the mature keratin complex.
Cells in the outermost layer of the stratum granulosum are dead and can no longer divide.
The cell death occurs due to rupture of lysosomes and the release lysosomal enzymes may complete the keratinization process.
The stratum granulosum is not uniform in distribution.
Especially in thin skin, it may be missing altogether.
It’s usually found in thick skin.
The stratum lucidum occurs only in non-hairy skin.
It consists of a layer of flattened keratinized cells that appears as a homogenous layer between the stratum granulosum and the stratum corneum.
The cells in this layer are dead and have a translucent appearance because they contain protein eleidin.
It’s the actual interface between the outer surface of the animal and its environment.
It’s the layer where cells are cast off in the normal process of wear and tear.
The stratum corneum consists of layers of fused, flattened cells devoid of organelles and filled with mature keratin.
The deep cells within this layer retain their desmosomal junctions.
Towards the surface, the desmosomes become disrupted and the cells become desquamated.
A group of cells that become loose and desquamated from the stratum corneum forms the stratum disjunctum.
Dermis ![]()
The dermis is a highly vascular dense fibroelastic connective tissue layer that contains blood vessels, lymphatics, nerves, hair follicles, sweat glands, sebaceous glands and many sensory receptors.
The dermis is divided into two layers: a superficial or papillary layer and a deep or reticular layer.
The papillary (superficial) layer is composed of a network of fine collagen, reticular and elastic fibers with fibrocytes, macrophages, plasma cells, mast cells.
Melanophores and fat cells may also be present in this layer.
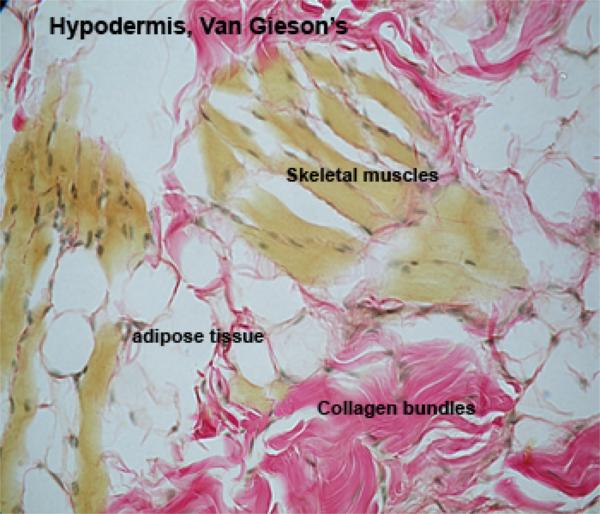
The hypodermis is a loose connective tissue layer containing variable amount of adipose tissue that attaches the dermis with the underlying tissues (bone or muscles).
Cells of the Epidermis
The epidermis contains at least four distinctive cell types: keratinocytes, melanocytes, Merkel cells and Langerhans cells.
They are the most numerous cell types in the epidermis.
They are concerned mainly with the production of keratin, hence the name keratinocytes.
These cells are produced by constant mitosis in the deeper regions.
With time they move up to the surface, die, and are worn away, to be replaced by the next wave of newly produced cells.
The keratinocytes begin as viable, dividing cells, and end as nothing more than flattened, battered, dead sacs of keratin.
Within the stratum basale, they are actively engaged in mitosis.
The cell migrates toward the stratum spinosum where they mature and accumulate tonofilaments and keratohyaline granules in their cytoplasm.
Within the stratum corneum, they are inactive and have no nuclei.
The tonofilaments and keratohyaline granules are compacted into flattened dead cornified cells.
The life of a given keratinocyte is about 7 to 10 days from production to sloughing.
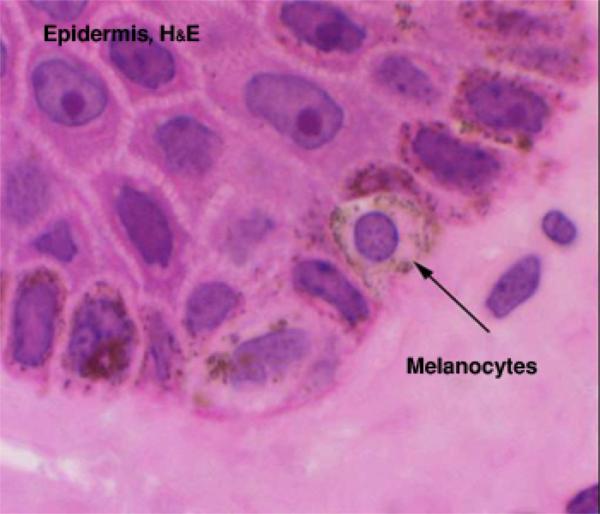
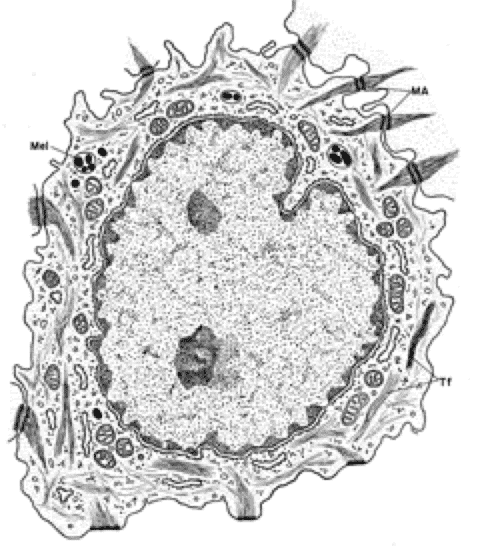
Melanocytes produce the protective pigment melanin. Melanin is responsible for skin and eyes color, and minimizes tissue damage by ultraviolet radiation.
Precursor cells that are destined to become melanocytes are derived from neural crest and migrate during prenatal life to the epidermal-dermal border where they differentiate into melanocytes.
With EM, the cells are pale-stained with large ovoid nucleus. The cell bodies of the melanocytes lie in the basal layer of keratinocytes in contact with the basement membrane. The cell bodies extend long cytoplasmic processes that extend between the keratinocytes.
The melanocytes produce melanin because they have the ability to synthesize the enzyme tyrosinase.
They tyrosinase is synthesized in rER and is transferred to Golgi where it is packed in a membranous vesicles known as premelanosomes.
Active tyrosinase within premelanosomes starts melanin synthesis, at this time the vesicles become known as melanosomes.
The melanosomes are ovoid membrane bounded granules that either remain widely dispersed as discrete structures or become aggregated into membrane-bounded melanosome complexes.
The melanoprotein complexes pass along the cytoplasmic processes of the melanocytes and are transferred into the cytoplasm of the basal and prickle cell layer keratinocytes.
Any cell type that phagocytizes melanosomes without having the ability to synthesize it is referred to as melanophores. Any type of melanin-containing particle that can be recognized by LM is described as melanin granule.
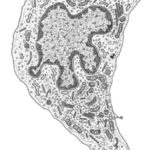
Merkel Cells ![]()
![]()
They are found in the basal layer of the epidermis.
The cells have cytoplasmic projections that indent or are insinuated between the keratinocytes to which they are anchored by desmosomes.
With EM, their cytoplasm contains electron-dense granules that are believed to synaptic vesicles.
The cells occur either as solitary cells or aggregates forming a structure known as tactile corpuscle.
The unmyelinated terminal branches of myelinated nerve fibers penetrate the basal layer of the epidermis, become expanded into a terminal disc that comes into contact with the base of the Merkel cells. Merkel cells are thought to be touch receptors.
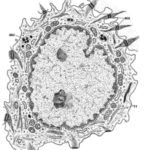
Diagram showing changes in epidermal cell shape during keratinisation and three specialised cells within the epidermis: m = melanocyte; L = Langerhans cell (of immune system); M = Merkel cell (associated with nerve ending).
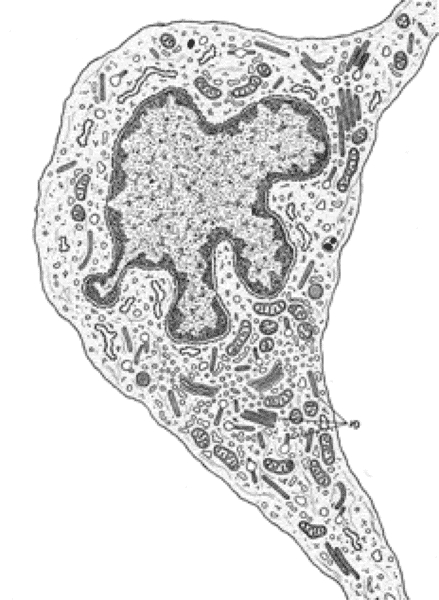
Langerhans Cells ![]()
They are bone marrow derived cells arrive by way of dermal blood vessels and then invade the epidermis.
Most Langerhans cells are situated in the stratum germinativum.
The cells are insinuated between keratinocytes but are not joined to them by cell junctions.
With EM, they can be distinguished from keratinocytes by their lack of tonofilaments (prekeratin filaments). Their nucleus generally exhibits irregular indentations.
Their cytoplasm contains a prominent Golgi complex, few rER, secondary lysosomes and moderate number of mitochondria.
The most distinctive feature of the cytoplasm is the presence of tennis racket-shaped
granules that are known as Langerhans cell granules or Birbeck granules.
The Langerhans cells serve as antigen presenting cells in the initiation of cutaneous contact hypersensitivity.
Differences between thin and thick skin
Thin skin differs from thick in a few ways, but mainly in the lack of a stratum lucidum.
The stratum granulosum is often absent or discontinuous.
The thin skin contains sweat glands, but they are less numerous than in thick skin.
The dermal papillae are lower, broader, and fewer in number than in thick skin.
Skin Appendages
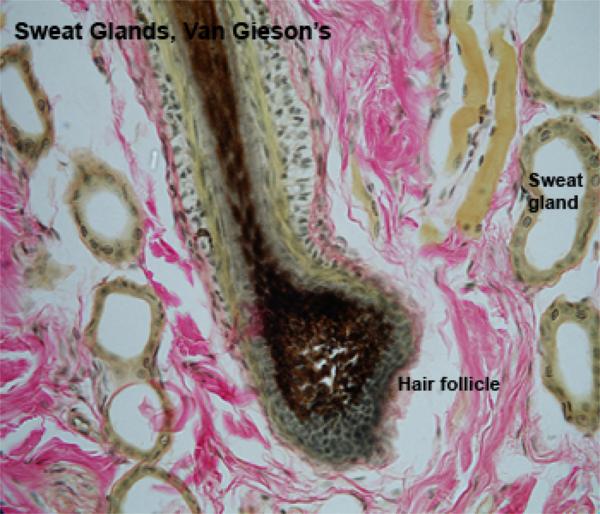
The skin appendages include hairs, hair follicles, sweat and sebaceous glands.
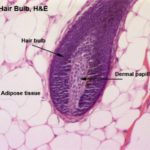
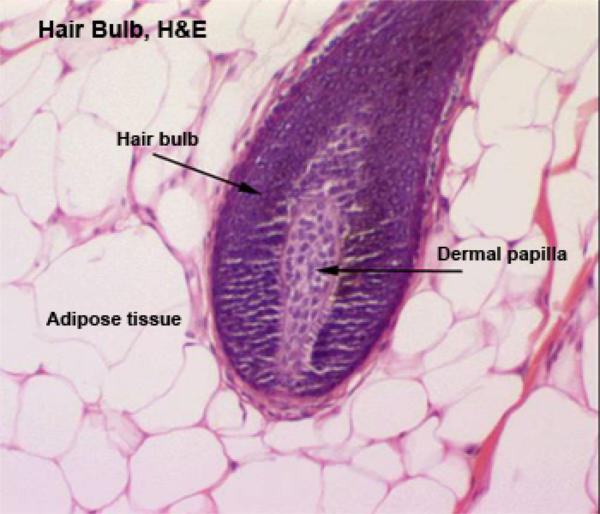
The hair is a flexible keratinized structure produced by a hair follicle. Each hair is composed of shaft or free part and root. The root of each hair has a terminal expansion known as the hair bulb.
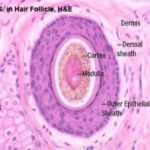
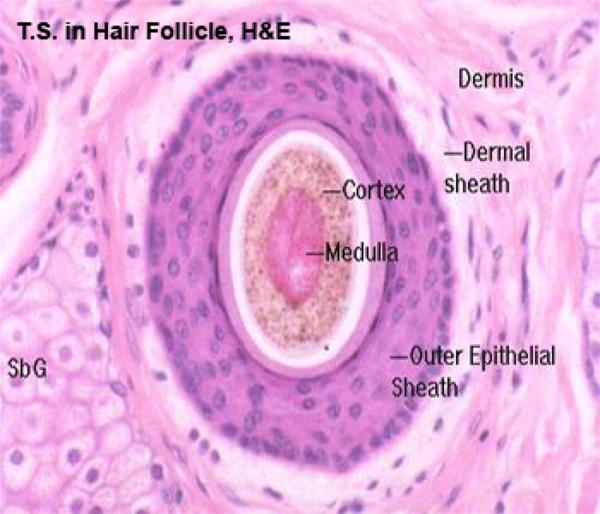
The hair bulb consists of actively dividing epithelial cells surrounding a papilla of connective tissue known as dermal papilla. The shaft of the hair is formed of three layers: cuticle, cortex and medulla.
The cuticle consists of is made up of a single layer of overlapping flat keratinized cells whose free edges are directed towards the distal (free) end of the shaft.
The cortex consists of a layer of dense, keratinized cells containing nuclear remnants and pigment granules. The medulla forms the center of hair and is loosely filled with cuboidal or flattened cells.
The hair follicle is a tubular epithelial structure opening onto the epidermal surface. Each hair follicle is formed of internal root sheath, external root sheath and connective tissue sheath.
Internal root sheath
The internal root sheath is made up of inner cuticle, middle granular layer and outer pale layer.
The inner cuticle consists of overlapping keratin plates and is continuous with the cuticle of the hair shaft.
The middle granular epithelial layer (Huxley’s layer) is formed of one to three layers of keratinized cells rich in large eosinophilic trichohyaline granules.
The outer pale epithelial layer (Henle’s layer) is composed of a single layer of keratinized cells.
The external root sheath
The external root is formed of several layers of cells similar to those of the stratum spinosum.
The connective tissue sheath
The connective tissue (dermal) sheath is a network of circular and longitudinal collagen and elastic fibers.
It is richly vascular and contains a delicate plexus of sensory nerve endings.
It is separated from the external root sheath by a thick basement membrane known as glassy membrane.
A bundle of smooth muscle called the arrector pili muscle is attached to the connective tissue sheath and is inserted into the dermal papillary zone.
Sinus or Tactile Hair Follicles
The sinus hair follicles are large single follicle characterized by a blood-filled sinus between the inner and outer layers of the dermal sheath.
In horse and ruminants, the sinuses are traversed by fibroelastic trabeculae. Skeletal muscles are attached to the outer sheath of the follicles allowing some voluntary control.
Numerous nerve bundles penetrate the outer sheath and ramify in the trabeculae and inner dermal sheath.
The sinus hair follicles are large single follicle characterized by a blood-filled sinus between the inner and outer layers of the dermal sheath.
In horse and ruminants, the sinuses are traversed by fibroelastic trabeculae. Skeletal muscles are attached to the outer sheath of the follicles allowing some voluntary control.
Numerous nerve bundles penetrate the outer sheath and ramify in the trabeculae and inner dermal sheath.
Merocrine Sweat Glands
The merocrine sweat glands of domestic animals are found mainly in special; skin areas such as footpads of the dog and planum nasolabiale.
They are coiled simple tubular glands that open directly onto the skin surface rather than into the hair follicles.
The secretory portion is composed of cuboidal epithelium with two distinct cell types.
The dark or mucoid cells have more ribosomes and numerous droplets in the apical part of the cell.
The clear cells lack cytoplasmic basophilia and they are believed to pump sodium ions into the gland lumen.
The duct is straight and composed of two layers of cuboidal epithelial cells resting on a basement membrane.
Myoepithelial cells form a discontinuous layer between the secretory cells and the basement membrane. Contraction of these cells expels sweat into the ducts.
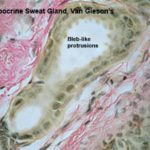
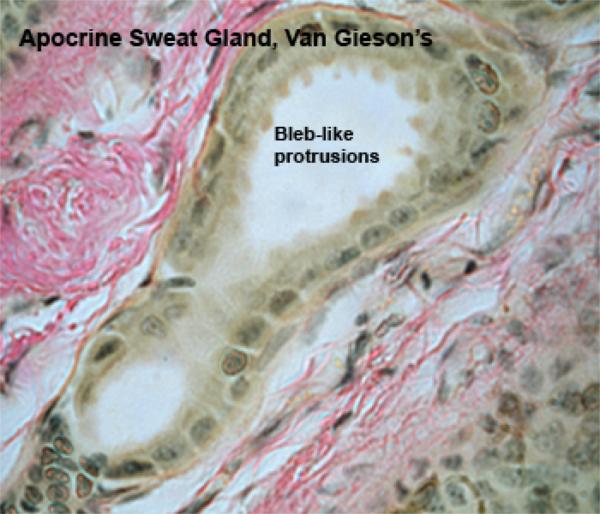
The apocrine sweat glands of the domestic animals are located throughut most of the skin.
In humen, they are found mainly in the axillary, pubic and perianal regions.
The glands are simple saccular or tubular with coiled secretory portion and a straight duct opening into adjacent hair follicles.
The secretory portion has a large lumen lined with flattened cuboidal to columnar epithelial cells.
The cytoplasm may contain glycogen, lipid or pigment granules.
The free surface of the cells has cytoplasmic protrusions. Myoepithelial cells are located between the secretory cells and the basement membrane.
The duct has a narow lumen and lined by two layers of flattened cuboidal cells.
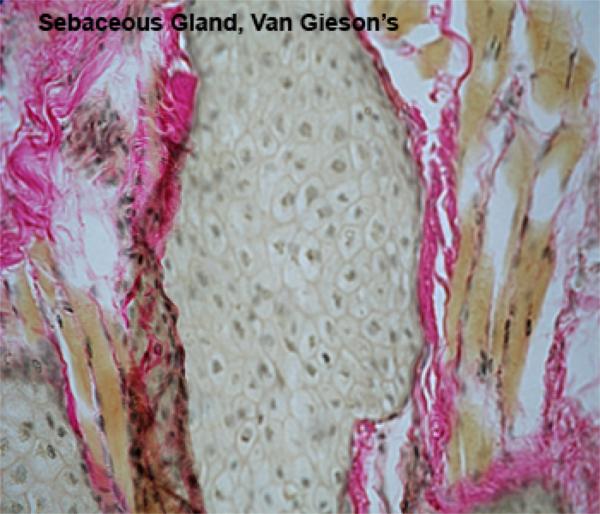
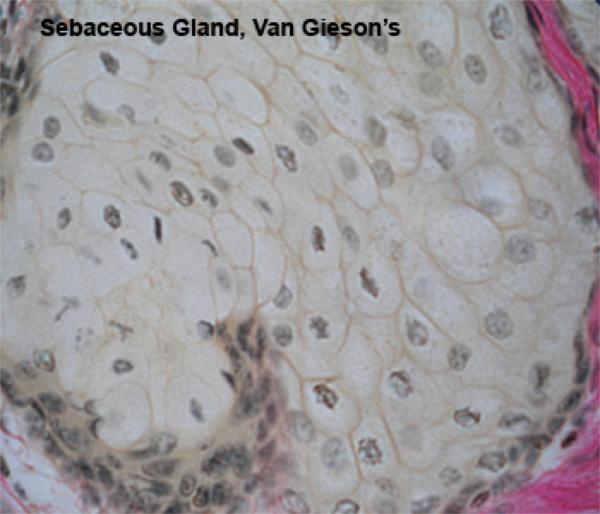
The sebaceous glands develop as lateral downgrwoths from the external root sheath. They are composed of lobules of large polyhedral pale-staining cells containing abundant lipid droplets and small dark staining central nuclei.
There is a single layer of cuboidal or flattened precursor cells between the basement membrane of each lobule and the central mass of cells.
The sebaceous glands are connected to the hair folllicle by short ducts lined by stratified squamous epithelium.
Towards the ducts, the lipid content of the cells increases greatly and the distended cells degenerate and release their contents into the duct (holocrine secretion).
Cells lost by holocrine secretion are replaced by mitosis in the basal layer of the acinus.