-
- Figure 1
-
- Figure 2
-
- Figure 3
-
- Figure 4
-
- Figure 5
-
- Figure 6
-
- Figure 7
-
- Figure 8
-
- Figure 9
-
- Figure 10
-
- Figure 11
-
- Figure 12
-
- Figure 13
-
- Figure 14
-
- Figure 15
-
- Figure 16
-
- Figure 17
-
- Figure 18
The male reproductive system consists of the testes, excurrent ducts, male accessory glands, urethra and penis.
The testes are exocrine and endocrine in nature.
They produce both a cellular secretory products in the form of sperm, and the hormone testosterone.
The testes of most mammals are located extra-abdominally in the scrotum.
This is actually a temperature-controlling device.
In most mammals spermatogenesis proceeds evenly and smoothly at an optimal temperature slightly below the core body levels.
The role of the scrotum is to maintain that temperature.
When the ambient temperature drops, the testes are pulled up by muscular action towards the warmth of the body cavity, and when it rises, the muscle relax, allowing the testes to descend and remain cool.
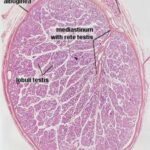
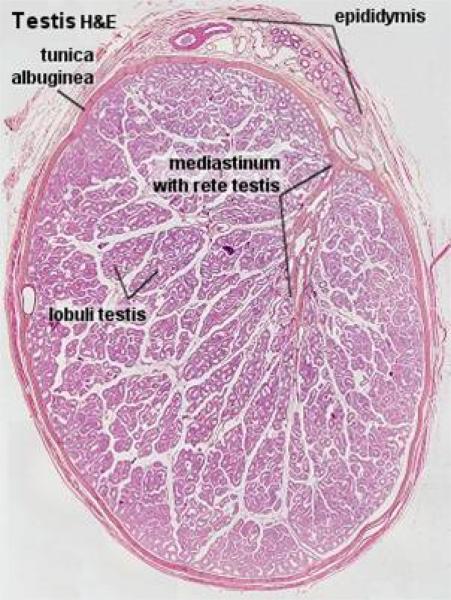
The stroma of the testis comprises a capsule, the tunica albuginea and the mediastinum testis.
The tunica albuginea (so named because of its white color on gross examination) consists of collagenous, few elastic fibers, numerous large blood vessels (stratum vascular) and smooth muscle (in horse).
From the tunica albuginea, small septa subdivide the cavity of the testis into smaller compartments.
Septation is usually incomplete in domestic animals except in carnivores.
The mediastinum testis is a central area of loose connective tissue that surrounds the rete testis and joins the septula testis which converge towards it from the tunica albuginea.
In most domestic animals, the mediastinum testis occupies almost the entire center of the testis.
In the stallion, the mediatinum testis is restricted to the cranial pole of the testis.
The testicular parenchyma consists of the seminiferous tubules, which are surrounded by delicate interstitial tissue.
The bulk of the testicular tissue is formed of the seminiferous tubules.
In human, the total length of the seminiferous tubules is about 600 meters, and the figure is much higher for large animals such as bulls and camels.
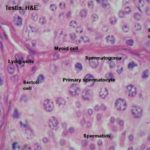
Each seminiferous tubule is bounded by a basement membrane surrounded by a single layer of boundary tissue cells similar to smooth muscle.
They’re sometimes called myoid cells or peritubular contractile cells, and at least in some species (e.g., rodents) they are known to be contractile, though it hasn’t been proven of all animals.
The contractions appear to be spontaneous, and not to involve any nervous stimulation.
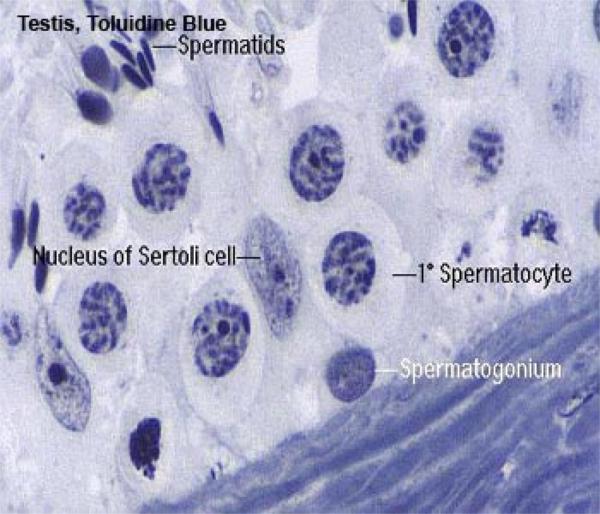
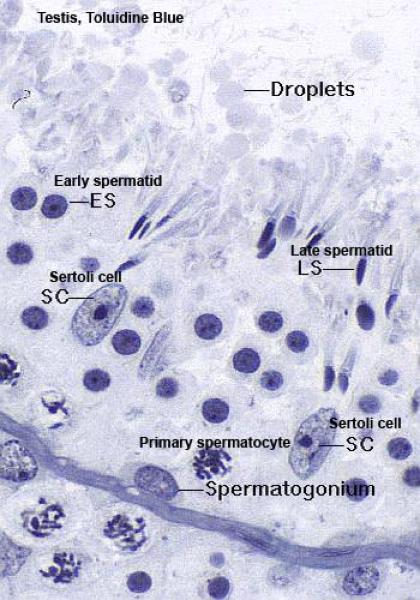
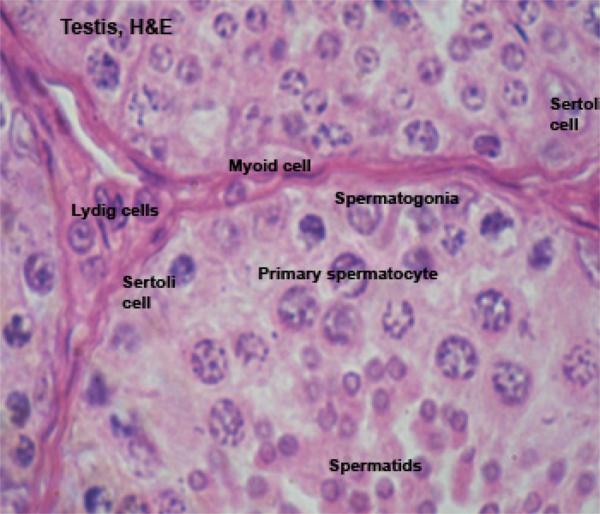
The seminiferous tubules are lined by seminiferous epithelium that comprises two cell types; Sertoli cells and spermatogenic cells.
The Sertoli cell or sustentacular cell is tall columnar or pyramidal with irregular outlines.
Each cell has a broad basal part rests on the basal lamina and a narrow apex extends into the lumen of the seminiferous tubules.
The cell is easily identifiable in H& E preparations by its nucleus.
The nucleus is pale staining, with an oval to pyramidal shape and one or two prominent nucleoli.
The cytoplasm is clear and poorly defined.
With EM, it contains abundant glycogen, lipid, sER, mitochondria, free ribosomes, and lysosomes.
![]() The spermatogenic cells are lodged in deep clefts within the lateral and apical surfaces of the Sertoli cells. The apical clefts contain the late spermatids.
The spermatogenic cells are lodged in deep clefts within the lateral and apical surfaces of the Sertoli cells. The apical clefts contain the late spermatids.
The Sertoli cells have several functions: 1) Protection and nutrition of the spermatogenic cells. 2) Phagocytosis of the residual bodies and damaged germ cells. 3) Secretion of the androgen-binding proteins. 4) Secretion of a small amount of
estrogen. 5) Formation of blood-testis barrier.
The haploid spermatocytes and spermatids are genetically different from the diploid spermatogonia.
Hence, there exists a possibility that the immune system might mount an immune reaction to them.
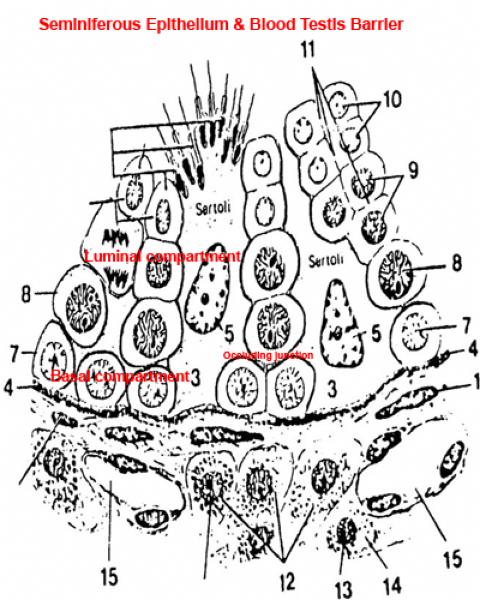
The cell’s lowest branches reach out to those of other sustentacular cells and fuse to form long occluding junctions.
The occluding junctions effectively separate the tubule into two compartments; one is below the fused lower processes (the adluminal or basal compartment), and the other is above them (the luminal compartment).
Blood borne components of the immune system are denied access to the haploid stages of sperm development by this blood-testis barrier.
As spermatocytes develop they need to move into the luminal compartment.
The appropriate sustentacular cells grow new processes, and the junctions above it are broken. The cell is now–without having passed through any membranes-no longer in the compartment accessible to the blood.
Spermatogenic cells ![]()
The spermatogenic cells include basal spermatogonia, primary spermatocytes, secondary spermatocytes, spermatids and spermatozoa.
Spermatogenesis
The spermatogenesis is the process by which spermatogonia divide and differentiate into spermatozoa. It can be divided into spermatocytogenesis and spermiogenesis.
Spermatocytogenesis
The spermatocytogenesis is the process of formation of spermatocytes from spermatogonia.
The spermatogonia are small cells of which there are two types designed A and B cells.
The spermatogonia A are cells with large spherical nuclei that contains fine dispersed chromatin and eccentrically situated nucleoli. Two types of spermatogonia A cell could be recognized; pale and dark. About half of the
spermatogonia A serve as stem cells.
The other half (probably the dark type) gradually differentiate over four successive mitotic division into spermatogonia B.
Spermatogonia B are spherical small cells with oval nuclei having coarse chromatin.
The cell divides by mitosis giving rise to primary spermatocytes.
The primary spermatocytes are the initial product of spermatogonial maturation.
They are the largest of the spermatogenic series and are located more centrally.
The cells have a long premeiotic prophase and a long life period of about 16 days in bull and 22 days in human.
The nucleus is coarsely granulated with visible chromosomes.
The nuclei are diploid but have double amount of DNA. The primary spermatocyte undergoes the first meiotic division giving rise to two secondary spermatocytes.
The secondary spermatocytes are about half the size of the primary spermatocytes.
They are located near the lumen of the seminiferous tubules.
They have haploid number of chromosomes and normal DNA amount.
They are difficult to be seen since they have a very short life span ranging from few minutes to an hour. They rapidly enter into second meiotic division giving rise to two spermatids.
N.B. Throughout the spermatogenic cell divisions, cytokineses is incomplete and the progeny of each cell are almost always interconnected by intercellular bridges
Spermiogenesis ![]()
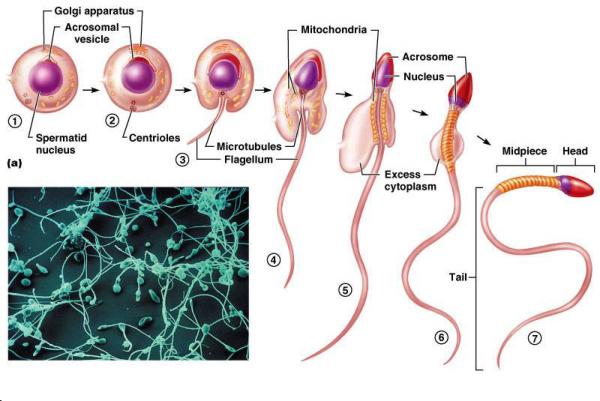
The Spermiogenesis is the process by which a spermatid is differentiated into spermatozoa. It comprises four phases, Golgi phase cap phase, acrosomal phase and maturation phase.
Golgi phase
The Golgi apparatus releases a large vesicle, the acrosomal vesicle.
The acrosmal vesicle attaches itself to the anterior pole of the nucleus.
One centriole migrates to the opposite pole of the nucleus and gives rise to a flagellum, the other migrates to form a cooler or annulus around the initial part of the tail.
Cap phase
The acrosomal vesicle flattens and spreads to cover the anterior half of the nucleus.
Acrosomal phase
The nucleus flattens and the acrosome covers its anterior two thirds. The region contains the centrioles forms the neck of the developing sperm.
As the flagellum elongates, nine coarse fibers become arranged longitudinally around the core of the flagellum.
The cytoplasm migrates to surround the first part of the flagellum.
The mitochondria become concentrated in the flagellar region and form a sheath around the coarse fibers.
Maturation phase
The excess cytoplasm is cast off from the spermatid in the form of a residual body that is phagocytised by the enveloping Sertoli cells.
Mature spermatozoon
The mature spermatozoon is formed of head, neck and tail.
The head contains the nucleus and is covered by the acrosome that contains hydrolytic enzymes.
The neck is relatively short and consists of centrally located centriole and nine peripheral longitudinally arranged coarse fibers continuous with the outer fibers of the middle piece.
The core of the middle pieces has a central axoneme (nine plus two arrangement of microtubules.(
The axoneme is surrounded by nine longitudinally arranged fibers that are further surrounded by a sheath of mitochondria.
The annulus marks the limit between the middle and principal piece.
The principle piece is the longest portion and consists of a central axoneme surrounded by inner and outer fibrous sheaths. The end-piece contains only an axoneme.
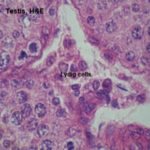
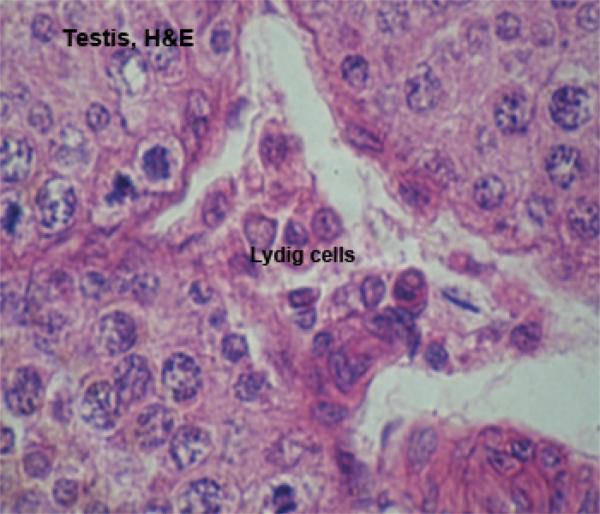
Interstitial cells (Leydig cells) ![]()
In the interstitial regions between the seminiferous tubules (i.e., outside of them) there are small groups of the cells which produce the male steroid hormone testosterone.
These are interstitial cells (or Leydig cells for Franz von Leydig, 1821-1908, a German anatomist).
They’re cuboidal or polygonal in shape with large spherical nuclei showing distinct nucleoli and present in-groups of 2-20 cells.
The cytoplasm is acidophilic vacuolated contains numerous lipid droplets, protein crystalloids (horse, cat) and pigment deposits, which increase with age.
With EM, the cytoplasm has abundant sER and mitochondria with tubular cristae (steroid-secreting cells).
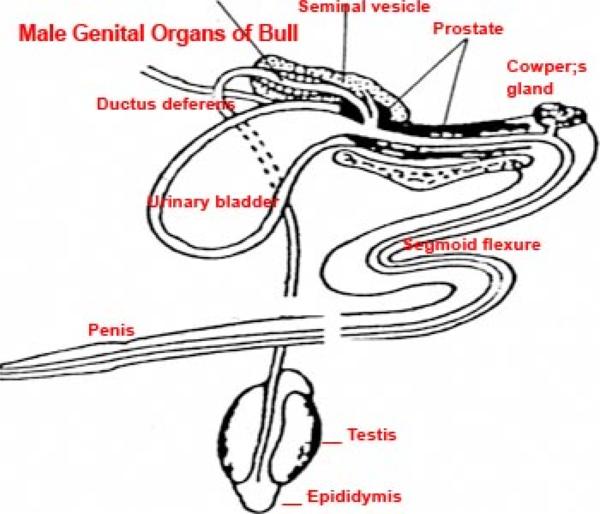
II. Male Excurrent Ducts
The male excurrent ducts conduct sperm to the outside and include tubuli recti, rete testis, efferent ductules, ductus epididymis, ductus deferens and male urethra.
Tubuli Recti
The tubuli recti connect the tubuli contorti with the rete testis.
They are lined by simple squamous to cuboidal epithelium.
The rete testis is the beginning of the outlet ducts.
It receives sperm directly from the seminiferous tubules and passes them on to the next portion of the drainage.
They are tortuous winding channels surrounded by the loose connective tissue of the mediastinum testis.
They are lined by cuboidal or stratified cuboidal epithelium.
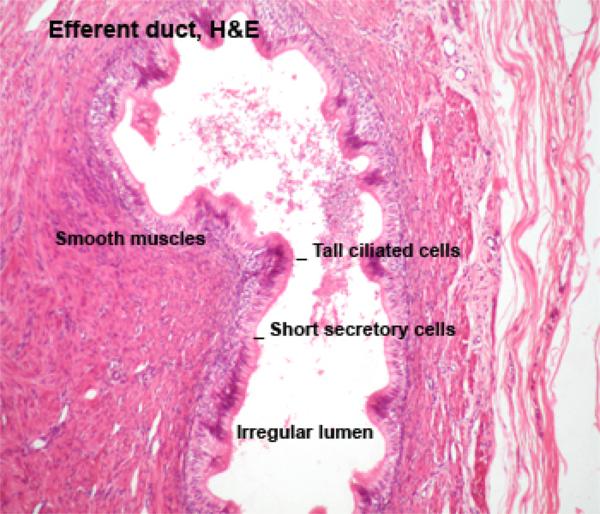
The rete testis opens into seven or eight efferent ducts.
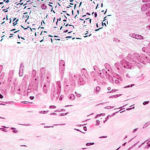
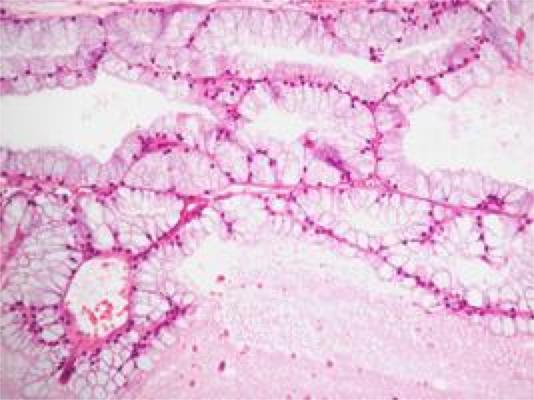
These coiled ducts are lined by a pseudostratified columnar epithelium of peculiar appearance, varying in height from place to place, giving the duct a scalloped appearance.
Many of the epithelial cells lining the duct, especially the taller ones, are ciliated.
These are true cilia, i.e., kinocilia, which have the typical internal structure and which are capable of movement.
The cilia help propel the not-yet-fully-matured and still immotile spermatozoa along their journey.
The efferent ductules penetrate the tunica albuginea and constitute together with the initial portion of the ductus epididymids the head of the epididymis.
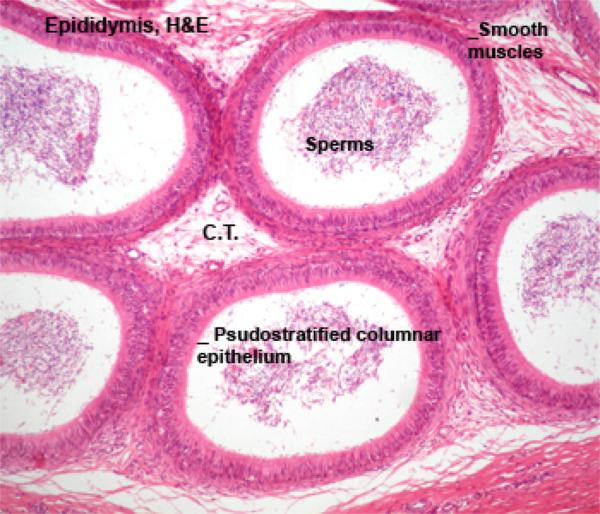
The epididymal duct is extremely long, highly coiled, originate from the fused efferent ductules.
It forms the body, tail, and a portion of the head of the epididymis.
The lining of the epididymis is pseudostratified columnar, but here they’re adorned with stereocilia rather than true cilia.
Stereocilia are unfortunately misnamed. They are not cilia at all, but instead are just enormously long microvilli.
They have no internal structure and do not move; they’re a specialization for increased surface area.
The small basal cells contain fat in ruminants and boar.
The epithelium contains vesicular intraepithelial glands (absent in ruminants).
The epithelium is surrounded by lamellae of connective tissue containing circular smooth muscle fibers.
The muscle layer is thin in the head of the epididymis, thicker in the body, and thickest in the tail region.
The epididymis serves for the slow transport, storage and maturation of the sperm, prior to ejaculation.
The epididymis is a secretory organ. It produces glycoproteins needed to coat the sperm and capacitate it, i.e., make it functionally capable of effecting fertilization.
Once the sperm have been resident in the epididymis for some time they are fully matured both morphologically and physiologically.
The epididymal cells have a very well developed Golgi apparatus, since they are actively involved in glycosylation reactions.
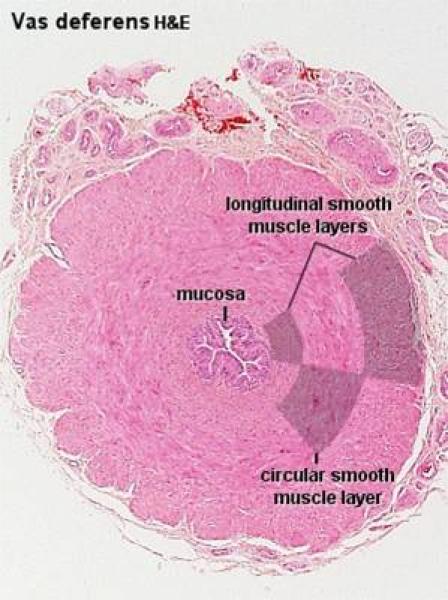
The wall of the ductus deferens is formed os mucosa-submucosa, tunuca muscularis and outer adventitia. The mucosa has longitudinal folds. The epithelium is simple columnar or pseudostratified columnar.
The propria-submucosa consists of loose connective tissue contains simple branched tubuloalveolar glands in the terminal portion.
The tunica muscularis is extremely thick, and is made up of inner and outer longitudinal and middle circular layer of smooth muscle.
The adventitia is a loose connective tissue layer rich is blood vessels and nerves.
In solipeds, ruminants and dogs, the terminal portion of the ductus deferens has a fusiform thickening forming ampulla ductus deferens. The propria-submucosa in the ampullary region is rich in simple branched tubular glands (ampullary glands),
which show sac like dilatations.
The gland tubules are lined by simple columnar secretory epithelium and their lumina often contain secretion.
In ruminants and horses, the lumina may contain crystals and calcified concretions.
The ampullary glands are absent in cat, small in boor, well developed in the solipeds and dog. In dog the glands extend into the initial portion of the urethra.
The Accessory Glands
The accessory glands of the male include ampullary glands, vesicular glands, prostate gland and bulbourethral glands.
All accessory glands are present in stallion, ruminants and boar, the vesicular glands are absent in carnivores, and the bulbourethral glands are absent in dog.
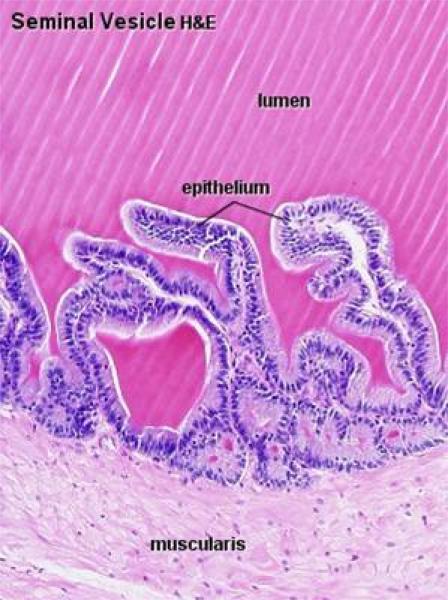
The vesicular gland or seminal vesicles are paired gland that are absent is carnivores.
The two seminal vesicles are tubular or turbuloalveolar glands. In solipeds and human, the seminal vesicles form true vesicular structure.
In ruminant and boar, they are lobulated glands.
The ruminants have large lobules with muscular septa. In boar, the lobules are small and the septa are fibrous.
The secretory tubules are surrounded by intralobular connective tissue rich in smooth muscle fibers.
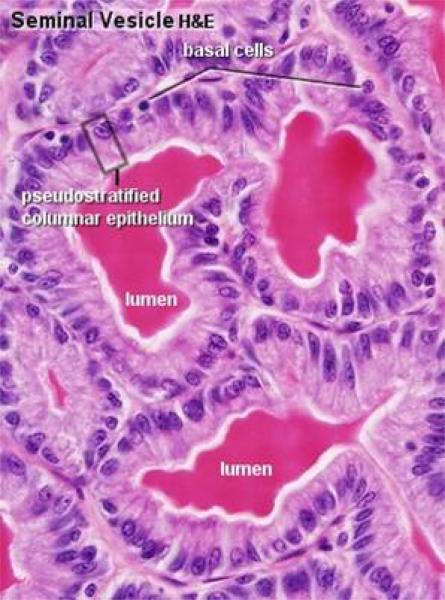
The glandular epithelium is pseudostratified columnar showing fat droplets in large ruminants.
In solipeds and human, the vesicular glands are true vesicles and their wall is formed of mucosa-submucosa, tunica muscularis and serosa.
The mucosa is folded. The epithelium is pseudostratified columnar.
The lamina propria-submucosa is a loose connective tissue layer contains sac-like secretory units lined by simple columnar secretory epithelium.
The tunica muscularis consists of longitudinal smooth muscle fibers.
The adventitia is a loose connective tissue outer layer. A portion of the gland may be covered by serosa.
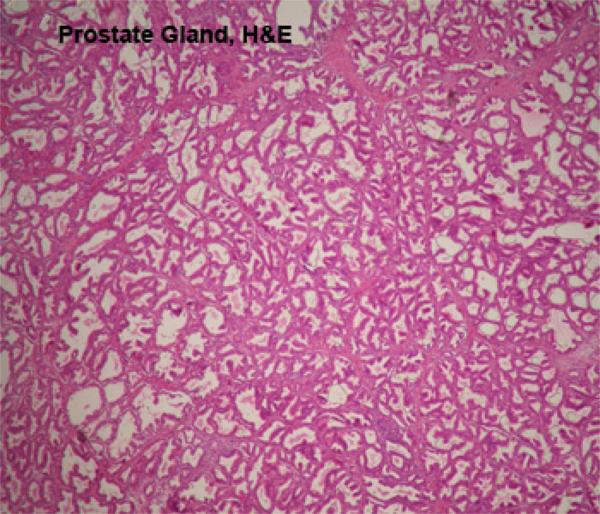
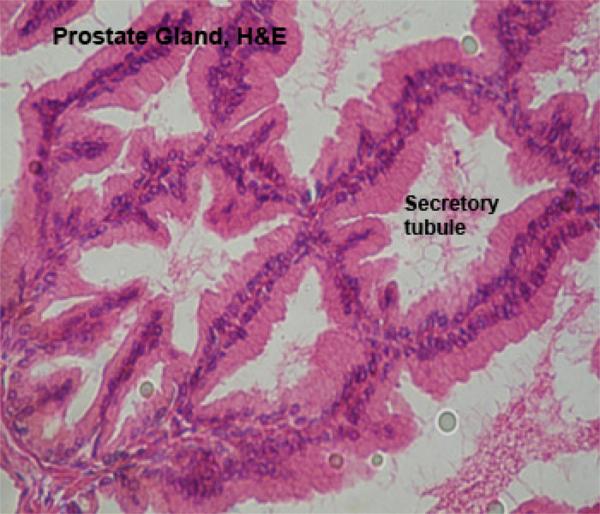
The prostate gland is a lobulated compound tubular gland located around the neck of the urinary bladder and is penetrated by the prostatic urethra.
The gland comprises of two portions; external portion (corpus prostate) which covers the pelvic urethra, and disseminated portion (pars disseminata prostate) which is located in the propria-submucosa of the prostatic urethra.
The gland is surrounded by a connective tissue capsule that sends septa dividing the gland into lobules.
The secretory tubules, alveoli and small ducts are lined by simple columnar epithelium.
In ruminants and swines, the pars disseminata is well developed and the corpus prostate is small.
In horse and carnivores, the corpus is much larger, and the pars dessiminata is represented by small glands (the urethral glands of Littre).
Bulbourethral Glands (Cowper’s Glands) ![]()
The bulbourethral glands are small, paired glands located on each side of the base of the penile urethra. They are absent in dog.
The gland is surrounded by connective tissue capsule containing a variable amount of smooth and striated muscle fibers.
Trabeculae of the same structure like the capsule extend to divide the gland into lobules.
The secretory tubules are lined by simple columnar secretory epithelium with bleb-like protrusion on their luminal surface.
The tubules drain into a central collecting duct that open into the urethra.
The Male Urethra
The male urethra is divided into pelvic and penile portions.
The pelvic urethra receives the secretions of all accessory glands.
The mucosa is folded and is projected at its dorsal part by the colliculus seminalis.
The epithelium is transitional. The propria-submucosa is a dense irregular connective tissue layer contains few cavernous blood spaces. In ruminants and boar, It contains the disseminated portion of the prostate gland forming a ring
of glandular tissue, which is thick dorsally, and thins, laterally and nearly absent ventrally.
The tunica muscular is formed of smooth muscle in the vicinity of the bladder and striated muscle in the rest of the urethra.
The adventitia is a loose connective tissue layer.
The uterus masculinus or prostatic utricle is an elongated vesicle lying in the colliculus seminalis. It represents the fused ends of the degenerated Mullelian ducts.
Penile Urethra
The penile urethra has fewer glands but more erectile tissue (corpus spongiosum) than the pelvic portion.
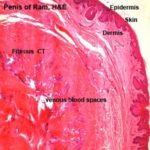
The cavernous tissue is composed of connective tissue trabeculae derived from the tunica albuginea that limits the erectile tissue.
The trabeculae contain many elastic fibers and blood vessels. Between the trabeculae there are many spaces lined by endothelium.
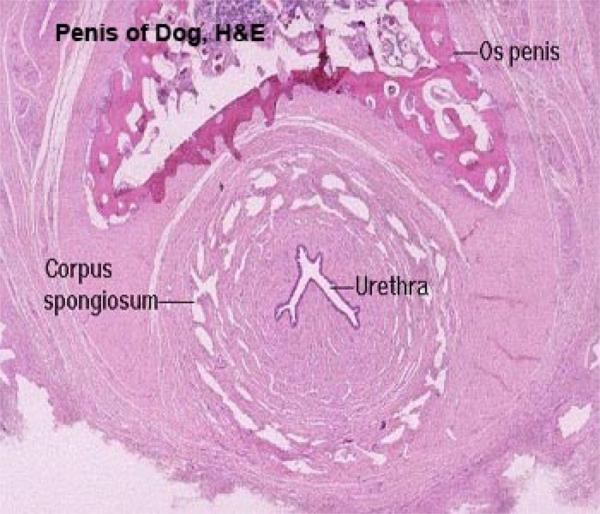
The body of the penis is formed of two dorsal corpora covernosa penis and a ventral corpus spongiosum penis surrounding the penile urethra.
The three corpora are enclosed by a dense fibrous connective tissue capsule, the tunica albuginea.
The tunica albugines sends septa into the erectile tissue.
Both the septa and the tunica albuginea contain elastic fibers and smooth muscle fiber.
A dorso-median septum (the septum penis) divides the corpora cavernosa penis either completely (dog) or only partially (other animals).
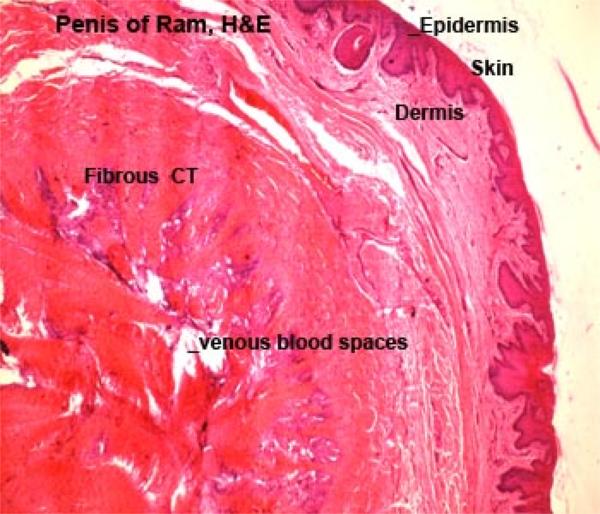
The spaces between the tunica albuginea and the trabeculae are filled with numerous venous (cavernous) spaces surrounded by smooth muscle cells and small amount of connective tissue.
In the stallion, the cavernous spaces are numerous and surrounded by scanty connective tissue, so the penis is classified as vascular or cavernous type.
In ruminants and boar, the caverns are scanty and the connective tissue is abundant, thus the penis is classified as fibroelostic type.
In dog and cat, their is a moderate amount of connective tissue and caverns, so the penis is classified as intermediate type.
Os penis, found in dog and cat. This triangular shaped bone is located dorsal to the urethra, and ventral to the corpus cavernosum, and its internal spaces are filled with marrow.
The glans penis is well developed in stallion and dog.
The outer surface is covered by skin. It is surrounded by tunica atbuginea that send trabeculae into the interior.
The spaces between the trabeculae contain cavernous spaces (erectile tissue).
The penis is covered externally by skin (visceral prepuce).
The penis of tomcat is similar to that of dog except, the spongy urethra courses along the dorsal surface. The visceral prepuce shows kornified papillae directed proximally.