-
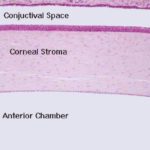
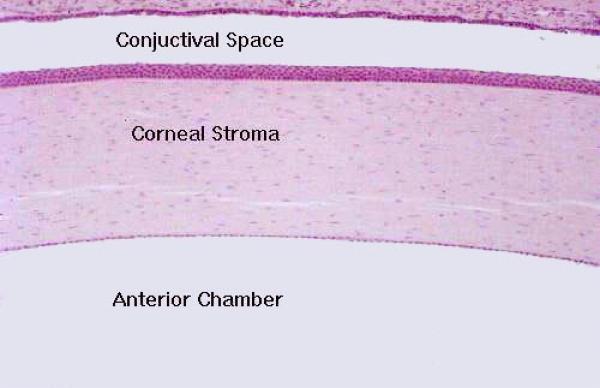
- Figure 1
-
- Figure 2
-
- Figure 3
-
- Figure 4
-
- Figure 5
-
- Figure 6
-
- Figure 7
-
- Figure 8
-
- Figure 9
-
- Figure 10
-
- Figure 11
-
- Figure 12
-
- Figure 13
-
- Figure 14
The Eye
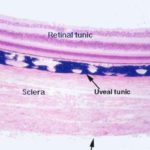
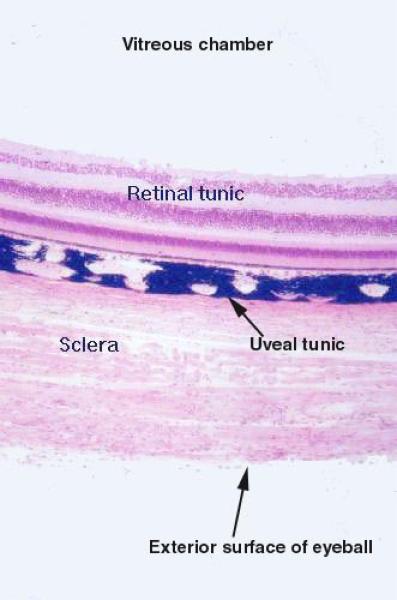
The wall of the eye is formed of three coats or layers:
Corneo-scleral tunic.
Uveal tunic or uveal tract.
Retinal tunic.
Corneo-scleral tunic:
The cornea is the thick, transparent portion of the coreo-scleral layer.
It forms the anterior one-sixth part of the corneoscleral tunic, joining the sclera at the limbus or corneoscleral junction.
The cornea is the principal light refracting structure.
The curvature of the cornea provides the principal mechanism for focusing images upon the retina.
Therefore, any alterations in the shape of the cornea can have profound effects on vision.
The cornea is exposed to the environment on its outer surface, and is an avascular structure consisting of five layers.
The outermost part of the cornea is a stratified squamous layer, the corneal epithelium.
The cells of the corneal epithelium sit on a thick basal lamina, Bowman’s membrane. The corneal epithelium varies in thickness among different species.
The bulk of the tissue in the cornea is the substantia propria, a thick collagenous connective tissue in the form of regular lamellae.
The inner surface of the cornea has a very thin, single layer of endothelial cells which are supported by a very thick elastic basement membrane known as Descemet’s membrane.
The cornea receives nutritional support by diffusion of nutrients from the aqueous humor and the blood vessels of the limbus; some oxygen is derived directly from the external environment.
Sclera ![]()
The largest part of the corneo-scleral tunics is the sclera.
This is what we call the “whites of the eyes,” and forms a flexible, watertight, and protective covering. It’s also the site of attachment of muscles that rotate the eye.
The sclera consists of dense, fibro-elastic connective tissue, the fibers of which are arranged in bundles parallel to the surface.
This layer contains little ground substance and few fibroblasts.
The scleral stroma is avascular.
The blood vessels and nerves running to and from the eye pass through the sclera.
Uveal tunic or uveal tract
The uveal tract is the middle tunic, and it’s composed of three elements.
These are the choroid, which comprises most of it, the ciliary body, and the iris.
Choroid
The choroid is the highly pigmented and vascular layer of the uveal tunic.
It provides nutritive support, and so there are numerous blood vessels and large lymphatic channels visible in it.
In most cases the melanin pigmentation in very obvious; the heavy pigmentation serves as a “light trap”.
It minimizes internal reflections and increases contrast by absorbing stray light passing through the retina, thus increasing the acuity of vision.
The chroid extends from the ora serrata to the optic nerve and it comprises chroid stroma, choriocapillary layer and Bruch’s membrane.
The chroid stroma is composed of loose fibrocollagenous tissue containing melanocytes, lymphocytes and mast cells, and through which the main arteries and veins run.
The choriocapillary layer is a capillary layer supporting the deep layers of the retina and arises from the larger vessels in the stroma.
The Bruch’s membrane is located at the interface between the chroid and the retinal pigmented epithelium.
It consists of the basement membrane of the choriopapillary endothelial cells, an outer collagen fiber layer, an elastic fiber layer, an inner collagen fiber layer and the basement membrane of the retinal pigmented epithelium.
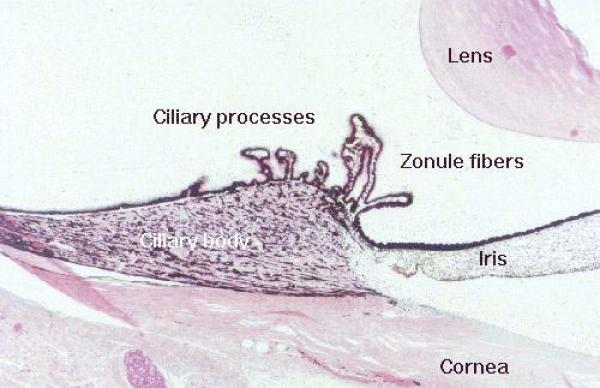
The ciliary body is continuous with the chroid and is located circumferentially between the ora serrata and the limbus.
Most of the ciliary body is composed of smooth muscle whose contractions work the lens accommodation mechanism via the tension imposed on the zonule fibers.
In addition to its role in the mechanism of accommodation, the ciliary body is a secretory structure.
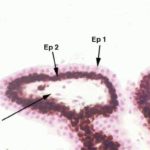
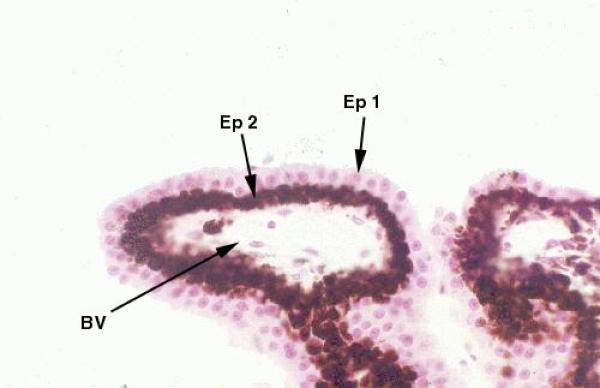
The ciliary body is lined by a dauble layer of cuboidal epithelium ![]() .
.
The outer of these two layers is heavily pigmented, the inner one is not.
This ciliary epithelium is the site of production of the fluid that fills the anterior and posterior chambers of the eye, the aqueous humor.
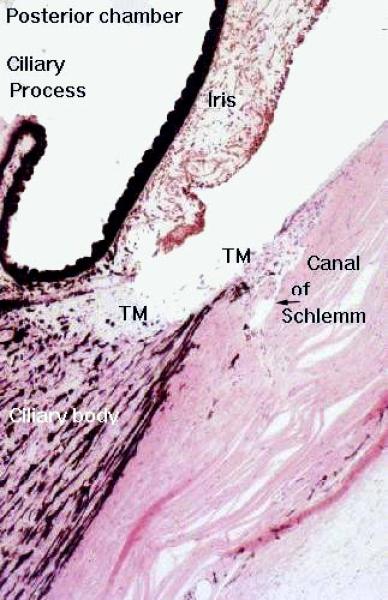
From the ciliary body finger-like projections known as ciliary processes extend into the posterior chamber.
The ciliary processes are responsible for the continuous production of aqueous humor, into the posterior chamber.
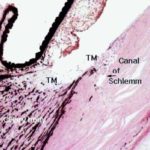
It then passes through the opening in the iris (the pupil) into the anterior chamber. Aqueous humor is then drained away via a series of channels in the angle between the iris and the cornea.
The principal site of drainage is the canal of Schlemm.
The canal is easily seen in the iridio-corneal angle as an irregularly shaped opening lined with simple squamous epithelium. It looks much like a small lymphatic duct or vein.
The continued production of the aqueous humor and its removal is important in function; the intraocular pressure thus produced helps to maintain the normal shape and size of the eyeball, which is vital to keeping an image in focus.
Blurry vision and “seeing stars” are premonitory symptoms of increased intraocular pressure, which can result from overproduction of aqueous humor, or damage to the drainage system, and this is the condition of glaucoma.
Uncorrected, pressure can build up and damage the eye, causing eventual blindness.
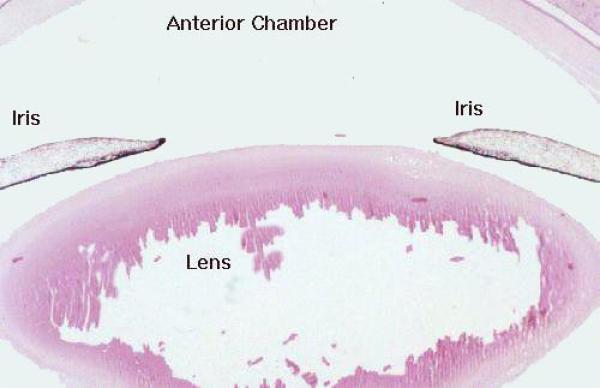
Projecting from the ciliary body is the iris. This forms a pigmented, diaphragm-like structure around the eye.
It has a hole of variable aperture in it, the pupil, through which light passes. The iris also serves to divide the eye into anterior and posterior chambers in front of the lens.
The main mass of the iris consists of loose, highly vascular connective tissue which is pigmented due to the presence of numerous melanocytes scattered in the stroma.
The anterior surface of the iris is irregular and consists of a discontinuous layer of fibroblasts and melanocytes.
The posterior surface is smooth and is lined by two layers of epithelial cells.
The surface layer is heavily pigmented while the deep layer is non-pigmented and is transformed into myoepithelial cells which constitute the radially oriented dilator pupillae muscle of the iris.
The constrictor muscle of the pupil (constrictor pupillae) consists of a band of circumferentially oriented smooth muscle fibers in the pupillary aspect of the stroma.
The muscle can contract in bright light or expand in dim light to provide the light sensitive retina with the proper level of illumination.
The iris forms the colored part of the eye.
The color of the iris depends on the amount of pigment in the connective tissue stroma. Blue eyes contain little stromal pigment whereas brown eyes have much stromal pigment.
The Lens ![]()
The lens is a biconvex, transparent structure composed of numerous layers of cells.
It’s located between the large vitreous chamber of the eye and the smaller posterior chamber.
It is composed of lens fibers that are extremely elongated epithelial cells that have lost their nuclei.
The cytoplasm of the lens fibers contains a crystalline protein and the cell membranes of adjacent fibers are fused leaving very little intervening extracellular substance.
The anterior aspect of the lens is marked by a single layer of nucleated cuboidal cells. The entire lens is enclosed by a homogenous capsule rich in glycoprotein.
The lens, because of its dense construction, is usually poorly infiltrated by paraffin, and histological sections usually show some damage and considerable shrinkage.
The lens is covered by a thin lens capsule, an amorphous layer that is really the basement membrane of the underlying lens epithelium.
The capsule is thickest on that side of the lens facing the iris, and thinnest on the posterior surface.
At the equator of the lens these epithelial cells proliferate, elongate, and transform themselves into lens cells.
The lens grows in size throughout life, and a constant production of new lens cells is necessary. Mitoses of the lens epithelium and differentiation provide for this.
The lens is responsible for accommodation i.e., the adjustment of the eye for observing near or far objects.
It does this by changing its shape.
his changes the focal length of the lens, and permits refraction of the rays coming from near or far objects.
The lens is suspended from the muscular ciliary body by a number of delicate zonule fibers.
Contraction of the muscles of the ciliary body tugs at the fibers and changes the shape of the lens.
In the relaxed condition (that is, with no tension on the fibers beyond that needed to support the lens) the shape is such as to refract rays from distant objects, and the eye is focused at infinity.
Accommodation for close vision requires tension to be exerted to deform the lens.
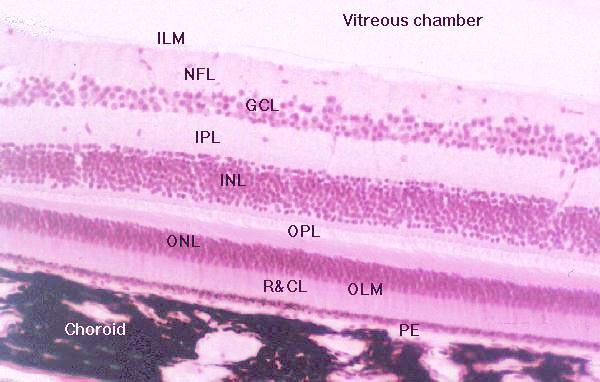
The retina is the innermost tunic of the eye, in which the light sensitive elements are located and on which the image is formed. Layers of the Retina
Pigment Layer
These cells comprise a layer of cuboidal cells that have spherical nuclei and an aggregation of melanin and lipofuscin at their inner ends.
The pigment layer cells phagocytose the ends of the rods and cones as they are renewed: hence the accumulation of lipofuscin.
The pigmentation helps to increase the contrast of the visual image by absorbing light that would otherwise be reflected back inwards towards the rods and cones.
Rods and Cones
The outermost layer of the retina proper is the bacillary layer or the layer of rods and cones. These are the actual light sensitive elements.
The rod photoreceptors are long, slender, bipolar cells.
The single dendrite of each cell extends beyond the outer limiting membrane as the rod proper.
The rod proper consists of inner and outer segments connected by a thin, eccentric strand of cytoplasm containing nine microtubule doublets similar to those of cilia but without the inner pair of microtubule.
The inner segment contains a prominent Golgi apparatus and many mitochondria. The outer segment has a regular, cylindrical shape and contains a stack of flattened membranous discs which contain the visual pigment rhodopsin.
Cone photoreceptors are similar to rod cells.
The outer segment is a long conical structure containing membranous discs that are continuous with the extracellular environment.
The cones contain pigment receptive to blue, green and red rather than rhodopsin. The bodies of the cone are continuous with the inner segment of the cone proper without an intervening dendritic process.
Rods and cones are transducers and their function is to take the physical signal of electromagnetic radiation and transform it into a neural one.
Rods are associated with low light vision, and are most numerous in nocturnal animals.
Cones are required for color vision (there are three types, one sensitive to each of the primary colors) and are most numerous in diurnal species.
Color vision is best in the birds and primates.
Outer Limiting Membrane
This isn’t really a membrane.
It’s really the site of numerous occluding junctions, which seal the rods and cones off from the inner portions of the retina.
This has the appearance of a membrane in the light microscope because there are so many junctions, all closely packed together.
The junctions are between the plasma membranes of the rod segments and a glial element, the Muller cell.
Outer Nuclear Layer
It is located immediately inward of the outer limiting membrane.
This layer is the location for the nuclei and cell bodies of the rod and cone cells. Collectively the nuclei of these cells constitute the outer nuclear layer.
Outer Plexiform Layer
The outer plexiform layer is the site of numerous synapses between the rod and cone cells and the processes of various integrator neurons.
While the initial neural impulse is generated in the rods and cones, they must pass the signal along to other neural elements for processing and further handling.
The dendrites of these cells and the axons of the rod and cone cells constitute the outer plexiform layer.
The integrating elements are the horizontal, bipolar, and amacrine cells.
They pass along the signal to the next neural element in the chain.
Inner Nuclear Layer
The cell bodies and nuclei of these integrator neurons are located in the inner nuclear layer, next inwards from the outer plexiform layer.
The bipolar cells are particularly important, as they have one pole extend outwards into the outer plexiform layer, and the other–their axon–extend inwards to the next layer, the inner plexiform layer.
Inner Plexiform Layer
The inner plexiform layer, like the outer one, is a region of synapses.
Here the bipolar cell processes synapse with the processes of ganglion cells. Ganglion cells are also neurons, and they are specifically those neurons whose axons will collect to form the optic nerve.
Thus they form the final intra-retinal element in the neuron chain, and their axons run back into the visual centers of the brain proper.
Ganglion Cell Layer
While the ganglion cells have synapses with bipolar cells in the inner plexiform layer, their cell bodies aren’t located there.
The cell bodies and nuclei of the ganglion cells are located in the next retinal layer, the ganglion cell layer, which has far fewer nuclei than the inner or outer nuclear layers.
Like any other neurons, these ganglion cells have axons, and the axons form the next layer inwards.
Nerve Fiber Layer
The axons of ganglion cells are bundled together to form the nerve fiber layer.
These axons are afferent fibers from the ganglion cells, and after running radially around the inner surface of the retina, they all come together at the site where the optic nerve penetrates the retina.
This is the “blind spot” of the vertebrate eye.
Inner Limiting Membrane
The innermost layer is the inner limiting membrane, which again isn’t really a membrane, but is instead a place of fused cell processes. In this case the fusion is between processes of the Muller ells.
Remember that most of the cells of the retina are in fact neural elements; they are either transducers (the rods and cones), integrators (bipolar and horizontal cells) or the terminal sensory neuron of the chain (the ganglion cells).
As is the case with other neurons, all of these cells are pretty much helpless to defend themselves against the insults of the outside world.
They have to be supported and coddled by neuroglia, or at least their equivalent.
The cells involved in the formation of the inner limiting membrane are Muller cells, tall cells which (like astrocytes in the CNS) serve to support and protect the neurons.
The Muller cells run from the outer limiting membrane to the inner limiting membrane, and are sealed at both ends.
At their outer ends they are fused to the “waists” of the rod and cone cells, and at the inner end to each other.
Thus the entire retina, with the exception of the expendable rod and cone cell outer segments (which, remember are renewed and phagocytosed) is completely sealed off from the blood vessels of the uvea at one end, and the vitreous body at the other.
The Optic Nerve and the Ora Serrata
The optic nerve begins at the optic disc, the point where all the axons of the ganglion cells come together, and projects back into the main portion of the brain.
This is the “blind spot,” of course, because no light sensitive elements can locate at the point of perforation.
There is another non-light sensitive portion of the retina that part between the ciliary body and the posterior part of the iris.
The margin between the sensitive and insensitive parts of the retina is the ora serrata, normally visible only in gross anatomy preparations.
Vertebrate eyes have a region of most acute vision, the fovea.
The retina is thinner here, and receptors more crowded together.
It’s an area where the thickness of the retina is reduced but the elements in it are noticeably more orderly and tightly packed.
The fovea is slightly off the optical center of the eye, and rays that come in at an angle to the central axis are focused directly on it. This arrangement gives an animal much better peripheral vision, which is of real survival value.
Accessory Structures
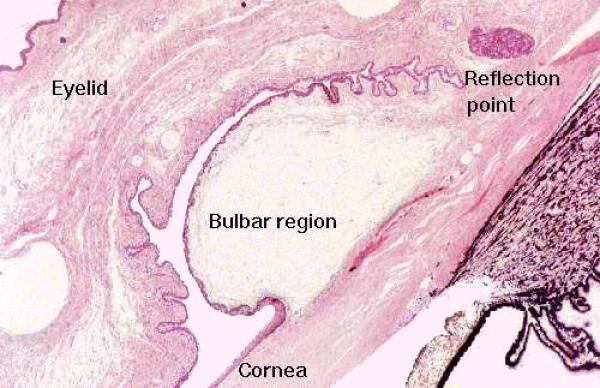
Accessory structures of the eye include the eyelids and the lachrymal glands.
The eyelid (and the part of the eye socket which faces it) is lined conjunctival epithelium.
The part lining the inside of the lid is the tarsal conjunctiva, which continues onto the outer surface of the eyeball as the bulbar conjunctiva.
The conjunctival epithelium is unusual in that it’s classified as stratified columnar, and it also contains goblet cells.
The tarsal conjunctiva forms a mucocutaneous junction with the integument at the edge of the eyelid, and the bulbar conjunctiva transitions to the stratified squamous epithelium of the cornea.
The center of the eyelid is filled with bundles of skeletal muscle, blood vessels, nerves, and glands.
The large tarsal glands (also called Meibomian glands are quite obvious.
These are modified sebaceous glands, opening into a long central duct.
There are also modified sweat glands present, the glands of Moll whose secretions are released into the follicles of the eyelashes.
It’s necessary to keep the surface of the eye moist.
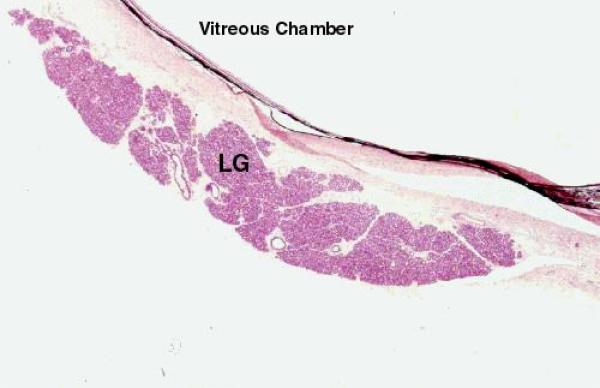
The eye produces a fluid secretion, the tears, by lachrymal glands.
Tears, in addition to providing lubrication, have an antibacterial effect because of the inclusion of lysozyme in their composition.
Ear
The ear is a complex organ with two functions: sound reception and maintenance of positional equilibrium.
Regions of The Ear
The ear has three distinct parts: the outer ear (the pinna and external auditory meatus); the middle ear (the tympanic cavity, the auditory ossicles, and the Eustachian tube); and the inner ear (the sealed, fluid filled chambers responsible for transduction of sound into nervous impulses and for providing information about body position).
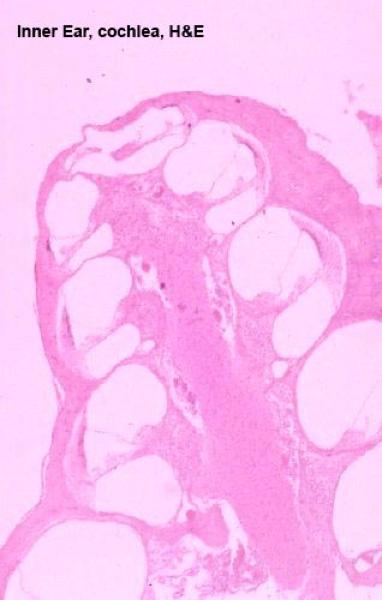
The Labyrinths of the Inner Ear ![]()
The inner ear is composed of bony labyrinth and membranous labyrinth.
The bony labyrinth consists of the convoluted channels in the actual bone of the skull. Inside these channels in bone sits the membranous labyrinth, components of which are the actual functional structures for balance and hearing.
The bony labyrinth is divisible into several regions: the vestibule, the semicircular canals and the cochlea.
The first two contain those parts of the membranous labyrinth that are involved in the balance sense.
The last part contains the portion of the membranous labyrinth that is involved in hearing perception.
Perilymphatic Spaces
The vestibule, cochlea, and semicircular ducts all are connected.
The vestibule is a fairly large space, and coming off it are the bony cochlea and the semicircular canals.
The cochlea is a corkscrew shaped passageway through the bone of the skull, and the semicircular ducts are simple loops that come off and return to the vestibule.
The bony labyrinth contains a small amount of fluid, the perilymph that fills it and in which the structures of the membranous labyrinth are bathed. Perilymph somewhat resembles extracellular fluid.
It acts as a shock buffer and a means of transmitting vibrations to parts of the membranous labyrinth.
The perilymphatic spaces are functionally connected to the subarachnoid space in the meningeal covering of the brain, and this may be the route by which excess perilymph is removed.
The actual site of perilymph production is a matter of debate.
Endolymphatic Spaces
The membranous labyrinth, too, is filled with fluid.
This fluid is endolymph, and it’s very different in composition from perilymph.
Endolymph is secreted by the cells making up the lining of the membranous labyrinth, and (unlike perilymph or tissue fluid) it’s high in potassium and low in sodium and soluble proteins.
The functional significance of the differences between these two fluids is not clear.
The Apparatus of Sound Transduction
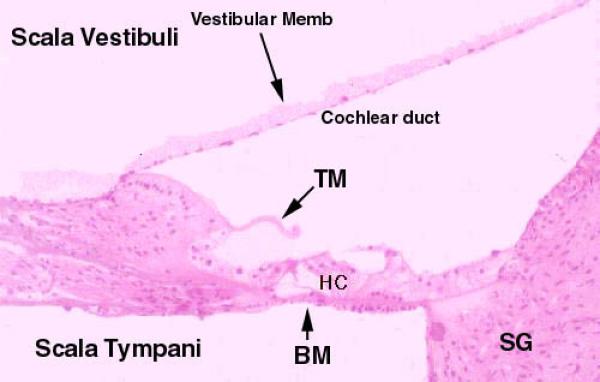
Cochlear Spiral: Scala Vestibuli, Scala Tympani, Scala Media
The cochlea is a spiral channel through the bone, and in the middle of it is a large bony projection which looks something like a wood screw, cut longitudinally.
This is the modiolus. Resting inside the cochlea is the cochlear duct, in which the events related to sound transduction occur. The duct follows the spiral of the cochlear lumen.
The cochlear duct is filled with endolymph, but the cochlea which surrounds is filled with perilymph.
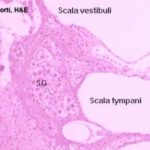
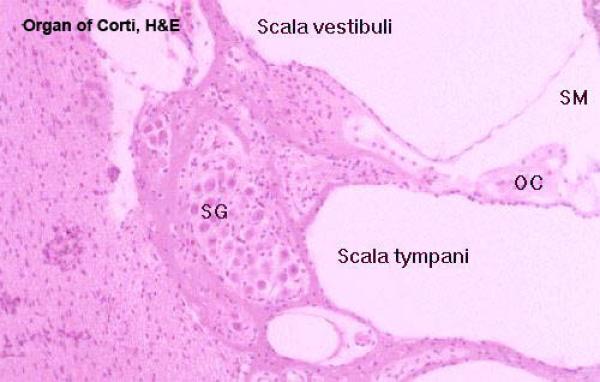
In cross section, each turn of the spiral shows three chambers; the first is the scala media, which is the cochlear duct itself, and in which are located the parts of the sound transduction apparatus.
Above the scala media, and separated from it by the very thin vestibular membrane is the scala vestibuli.
Below the scala media, and separated from it by the thick basilar membrane, is the scala tympani.
The scala tympani is also a perilymphatic space, is also in communication with the vestibule, and is also in communication with the scala vestibuli.
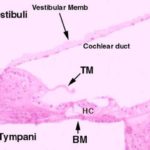
Scala Media and Organ of Corti ![]()
![]()
Inside the cochlear duct (scala media) itself is the actual organ of sound transduction. This is the organ of Corti.
The organ of Corti is a long, flat, spiral structure which follows the turns of the cochlear duct as it winds up to the top of the cochlear spiral.
When the organ of Corti is viewed in cross section it appears as two groups of cells sitting on the basilar membrane; these are surmounted by a thin, transparent, and acellular tectorial membrane.
The tectorial membrane is made of a keratin-like material, and in H&E sections rather resembles a piece of fingernail.
It’s anchored at one end to the wall of the cochlear duct, and it’s quite stiff. It, too, is spiraled and it covers the two groups of cells all the way up to the top of the spiraling scala media.
The hair cells are the actual transducers of mechanical movement into neural impulses.
Each is an epithelioid cell with projecting “hairs” that contact the bottom of the overlying tectorial membrane.
It’s important to realize that these aren’t “hairs” at all, of course; they are microvilli and modified cilia.
It should be fairly obvious that any movement of the flexible basilar membrane will deform the hair cell projections against the tectorial membrane.
The hair cells are transducers, specialized detector elements, and they’re surrounded by sensory fibers of neurons whose cell bodies are in the spiral ganglion.
Deformation of the hair cell projections causes changes in their membrane potentials, which are detected by the cells of the spiral ganglion.
Thus, mechanical movement of the basilar membrane relative to the fixed tectorial membrane results in the generation of a nervous signal.
Mechanics of Sound Detection
Sound waves coming in via the external auditory meatus cause the eardrum to move back and forth.
In the middle ear, a series of small bones constituting a lever system magnifies this initial signal and mechanically transmits it to an opening in the side of the vestibule.
The pulsations of this movement are then sent into the perilymph as actual fluid waves.
The waves induced in the perilymph of the vestibule travel UP the scala vestibuli to the top of the cochlea.
Since the scala tympani and the scala vestibuli are in communication at the top of the spiral (the term for this area is the helicotrema), the fluid pressure wave in the perilymph “crosses over” and starts back DOWN again, via the scala tympani.
Hence, there is perilymphatic fluid moving on both sides of the scala media.
All this sloshing back and forth in the perilymph causes the suspended cochlear duct (which is filled with endolymph) to vibrate as well.
This moves the basilar membrane up and down, causing scraping of the hair cells along the bottom of the tectorial membrane.
The hair cells send the resultant signal out via the cells of the spiral ganglion.
The cochlear nerve consists of the axonal projections of the cell bodies in the spiral ganglion and, like any other sensory nerve, runs back into the brain. In this case, it becomes one of the components of cranial nerve VIII, the vestibulocochlear nerve.
Balance Sensation
The Vestibular Apparatus
The vestibuleis the portion of the bony labyrinth from which the cochlea and the semicircular canals arise.
As with other bony labyrinth, the vestibule and the canals are filled with perilymph, and in them are suspended parts of the membranous labyrinth, constituting the vestibular apparatus of balance sensation.
As is the case with the cochlear duct, the vestibular apparatus is filled with endolymph and sealed off from the surrounding perilymphatic spaces.
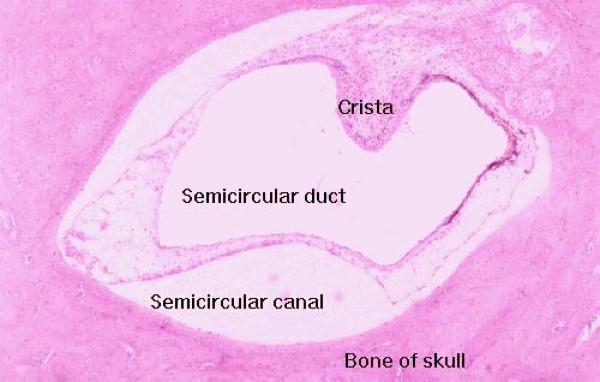
The vestibular apparatus consists of three large dilated areas and three semicircular ducts which run through channels of the bony labyrinth.
The semicircular ducts (and the semicircular canals through which they run) are oriented in three planes of movement: vertical, horizontal, and anterior-posterior.
Inside the semicircular ducts are elevated areas of epithelium, covered with hair cells similar to those of the organ of Corti.
These are the cristae ampullares, they project out into the lumen of the semicircular duct.
It’s covered with a pseudostratified epithelium composed of hair cells and some supporting cell types.
The hair cells, like those of the sound detection system, are transducer cells with deformable elements on the free surface.
At their bases, they are surrounded by the ends of nerve fibers from the vestibular branch of the vestibulocochlear nerve.
The hairs in this organ are polarized–that is, they are of different lengths, so that there is a directionality in their arrangement with respect to the axis of the semicircular duct.
Movement of the fluid in the semicircular duct will obviously cause deformation of the hairs, and this in turn will cause depolarization of the hair cells and a change in their membrane potential.
This change is transmitted via the sensory fibers and the vestibulocochlear nerve to the brain.
This is the same sort of mechanism at work in hearing; mechanical movement of fluid is translated into neural impulses.
Movement of the head in any direction causes the fluid in the semicircular ducts to flow relative to the hairs, and sends the sensation of movement to the brain.
The vestibular branch of the nerve joins the cochlear branch to form the vestibulocochlear nerve coming from the spiral ganglion.
Function of the System
This system is designed to detect acceleration–i.e., changes in speed or direction over a given time.
Any movement of the head relative to the three planes in which the semicircular ducts are fixed produces a corresponding movement of the endolymph inside them.
This deforms the hairs and provides information to the brain about the direction of movement.
Gravity detection
There is one other portion of the vestibular system This is the maculae.
The maculae are located in the large bulbous expansions of the vestibular system (also filled with endolymph), and their function is to provide a reference to the pull of gravity.
The expanded portions of the membranous labyrinth are the saccule and the utricle.
Both these regions contain areas with hair cells similar to those in the cristae.
Unlike the semicircular duct system, the sensory areas of the maculae don’t depend on fluid movement.
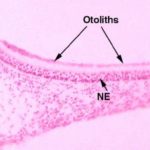
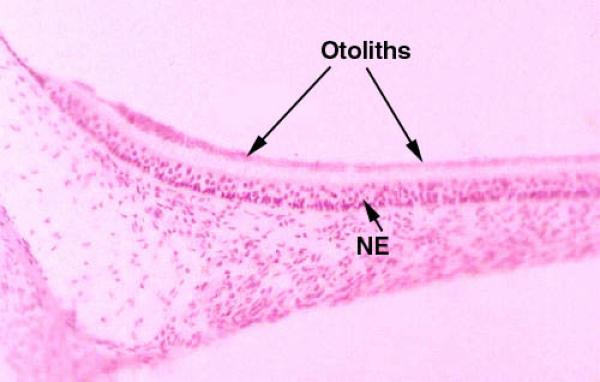
The maculae have within them small calcareous secretions, the otoliths ![]() , or “ear stones,” which lie on top of the hairs of the hair cells.
, or “ear stones,” which lie on top of the hairs of the hair cells.
Otoliths are formed in the embryo as part of normal development.
When an animal is stable with respect to gravity, the force the otoliths exert on the hairs is more or less constant, and the brain interprets this as “I am not moving downward, I’m sitting still on the ground.”
If, however, the animal falls, the otoliths exert different forces on the hairs, causing a different input to the CNS.
Otoliths in many non-mammals and invertebrates often contain iron and are affected by magnets.
If you tape a magnet to the shell of a crab, for example, his otoliths are attracted to it, and he’ll swim upside down.
Some migratory birds make use of the magnetic field of the earth for navigation, and have a built in compass in their otoliths.
Reports indicate that taping magnets to the heads of pigeons and migratory waterfowl causes them to lose their way.